| Publié par (l.peltier) le 31 décembre 2008 | En savoir plus |
2 milliards
Une grosse bactérie capte deux minuscules bactéries, peut-être tout simplement pour s’en nourrir. L’une est une algue bleue capable d’effectuer la photosynthèse ; l’autre, une bactérie [1] performante du point de vue énergétique. Le résultat de cette symbiose est la cellule végétale telle que nous la connaissons aujourd’hui, équipée de ces deux symbiotes [2] présumés : le chloroplaste [3] vert et la mitochondrie [4] énergétique. Il n’existe aujourd’hui aucun type intermédiaire entre les cellules bactériennes, les protocaryotes, et les cellules complètes pourvues d’un noyau, de chloroplastes et de mitochondries, dites eucaryotes.
Jean Marie Pelt. La solidarité chez les plantes, les animaux, les humains. Fayard 2004
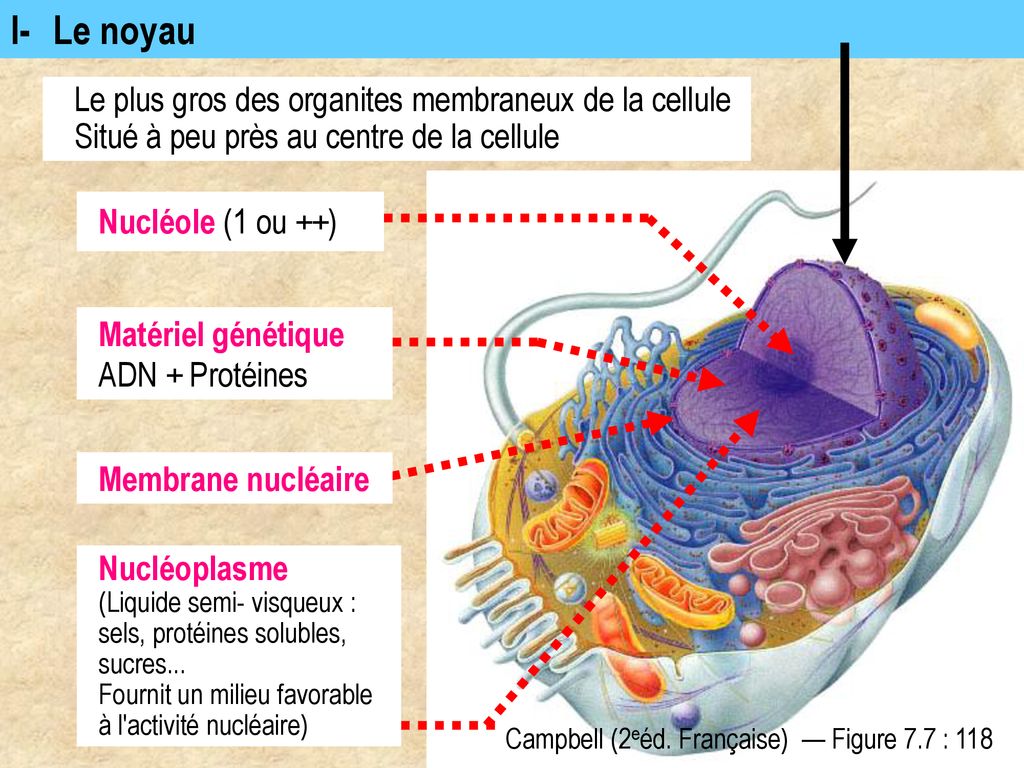
La cellule eucaryote se reproduit par mitose et ou méiose ; parmi les organites qui la composent, outre les mitochondries, on a encore le reticulum, le dictyosomer et plastes chez les végétaux ; elle est pourvue d’une paroi cellulaire pectocellulosique et elles ont un cytosquelette (actine, microtubule)
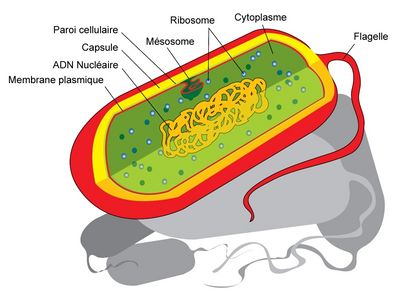
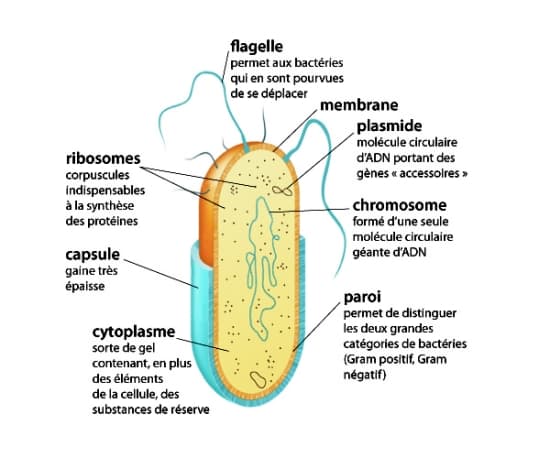
Bactérie
L’apparition d’un pigment particulier, la chlorophylle, entraîne une véritable révolution provoquant une inévitable mutation du monde végétal, mais aussi des changements climatiques et des modifications de la composition de la croûte terrestre. Le processus auquel ce pigment est associé s’appelle photosynthèse. Les premiers organismes capables de réaliser la photosynthèse – la plante absorbe du gaz carbonique, fabrique des sucres et libère de l’oxygène – utilisaient donc l’énergie solaire, ils étaient en mesure de produire une quantité d’énergie dix fois supérieure à celle qui pouvait être obtenue par fermentation, mais cette énergie pouvait être stockée comme réserve énergétique sous la forme d’un composé carboné, le glucose. Ces organismes dépendaient seulement de la disponibilité d’eau et de gaz carbonique. Certains dépôts très anciens appelés stromatolites témoignent de l’activité très importante de ces organismes unicellulaires.
Le début de la photosynthèse signifie l’apparition, à dose conséquente dans l’atmosphère, d’oxygène, véritable poison pour le monde vivant de l’époque. Le taux dans la composition de l’atmosphère va atteindre progressivement le taux actuel de 18 %, obligeant les organismes vivants à évoluer pour survivre. Les oxydes de fer sont les témoins de cette augmentation du taux d’oxygène, et les principaux acteurs des changements de la composition de la croûte terrestre. Dans la haute atmosphère, le dioxygène O² se transforme en trioxygène O³ – l’ozone -. Une couche d’ozone, cela signifie que les ultra-violets sont filtrés et que les cellules végétales, jusqu’alors brûlées, peuvent désormais continuer à vivre.
La Terre s’est formée il y a 4,6 milliards d’années. Son atmosphère était composée de vapeur d’eau et de gaz carbonique. Après refroidissement, la vapeur d’eau s’est condensée en eau liquide, inondant la croûte terrestre et engendrant les océans. Le gaz carbonique s’est dissout dans l’eau, et en présence d’ions calcium (Ca++) a formé du calcaire (CaCO³).
La vie dans la mer est apparue il y a 3,8 milliards d’années sous forme de bactéries. Cette vie était sans oxygène : anaérobie.
Ces bactéries possédaient certes des systèmes biochimiques d’oxydo-réduction, mais leur potentiel n’était pas suffisant pour oxyder l’eau en oxygène et hydrogène grâce à l’énergie des photons du soleil.
C’est alors qu’apparut chez certaines bactéries il y a 2,8 milliards d’années, une nouvelle molécule biochimique : la chlorophylle. Ces bactéries étaient et sont toujours des cyanobactéries appelées aussi algues bleues.
La chlorophylle de ces organismes unicellulaires captant l’énergie solaire va permettre, par son potentiel d’oxydo-réduction élevé, d’oxyder l’eau (H²O) en Oxygène (O²) et Hydrogène (H²). Grâce toujours à la molécule de chlorophylle, les cyanobactéries vont réaliser une photosynthèse en utilisant l’énergie solaire pour transformer le gaz carbonique et l’hydrogène en molécules organiques. Ces cyanobactéries sont dites autotrophes, c’est-à-dire capables de transformer des substances minérales CO2 et H2 en matière organique pour vivre. L’énergie lumineuse se trouve convertie en énergie chimique dans les maisons chimiques constituant les molécules organiques.
La vie était cantonnée dans l’eau. Mais l’oxygène produit au cours de l’oxydation de l’eau n’était pas utilisé, c’était un déchet.
C’est à cette époque, à cause de l’oxygène non utilisé, que la plus grande catastrophe écologique de tous les temps se produisit.
L’oxygène est transformé en ion superoxyde, radical libre possédant un électron célibataire, doué de propriétés destructives de molécules organiques en rompant des liaisons chimiques. Un poison d’une extrême toxicité a envahi la mer.
Mais des bactéries anaérobies, rescapées de cet holocauste, se mirent à développer un système de défense : la chaîne d’oxydo-réduction phosphorylante des cytochromes : la respiration naquit.
Les molécules organiques synthétisées à partir du gaz carbonique et de l’hydrogène sont métabolisées en CO2 et libèrent des H2. Ces H2 vont être scindés en protons et électrons 2H+ et 2e-. Ces électrons vont s’écouler sur des systèmes d’oxydo-réduction de potentiel décroissant jusqu’à l’oxygène 1/2 O2 qu’ils réduiront en O–.
Les protons 2H+ libérés formeront avec O– une molécule d’eau H²O.
La majorité des molécules d’oxygène est neutralisée, la toxicité de l’oxygène par le biais de la production de radicaux libres disparaît : la vie aérobie peut avoir lieu.
De plus l’écoulement des électrons n’est pas comme une rivière tranquille, il y a des chutes d’eau qui sont ici des chutes de potentiel d’oxydo-réduction.
À chaque chute de potentiel d’oxydo-réduction une énergie sera libérée et emmagasinée dans une molécule d’ATP Adénosine Tri Phosphate.
L’homme est un être dont l’organisme est pluricellulaire. Il ne possède pas de chlorophylle et doit puiser son énergie à partir de l’oxygène qu’il respire et des aliments qu’il consomme. Ces aliments sont des glucides, des lipides et des protides. Au cours de la digestion ces constituants seront dégradés en glucose, acides gras, acides aminés et seront catabolisés dans le cytoplasme cellulaire en CO2 et en dérivés d’hydrogène AH2. Le CO2 sera expiré et passera dans l’atmosphère. AH2 se dissociera dans les mitochondries en A et H2 et ensuite H2 donnera 2H+ et 2 e-. Ces mitochondries dont les ancêtres étaient des bactéries possèdent la chaîne des cytochromes d’oxydo-réduction phosphorylante. Le bilan de cette chaîne respiratoire est la réduction de l’oxygène Vz 02 + 2e– = O– et la production d’énergie sous forme d’ATP.
L’oxygène réduit O- avec les 2 H+ formera de l’eau H2O. L’ATP hydrolyse en ADP et P04—et fournira par rupture de la liaison (riche en énergie) l’énergie nécessaire pour la vie, énergie reconvertie en énergie chimique, cinétique, thermique, électrique,
En plus pour que la vie apparaisse sur la terre ferme il fallait que l’intensité du rayonnement solaire riche en ultra-violet diminue. Une barrière protectrice s’est mise en place grâce à l’oxygène. Dans la stratosphère, des molécules d’oxygène se dissocièrent et les atomes d’oxygène libérés (O) se combinèrent à des molécules d’oxygène (O2) pour former de l’ozone O3.
Cette couche d’ozone va progressivement s’épaissir et permettre par absorption des rayonnements U.V.
Si la majorité des molécules d’oxygène sont, par la respiration transformées, en ions O—, il demeure que des molécules d’oxygène peuvent dans les mitochondries, au cours de certaines circonstances (mode de vie, environnement) évoluer en radicaux libres sous forme d’ions superoxyde, de radical hydroxyl.
Ces radicaux libres vont alors s’attaquer aux constituants vitaux de nos cellules.
Ils peuvent rompre des liaisons chimiques de l’ADN, perturbant sa réplication et entraînant mutations et cancers. Ils peuvent également oxyder les protéines et les lipides des membranes cellulaires.
Heureusement ces actions sont partiellement contrées par l’élaboration d’enzymes capables de neutraliser ces dérivés toxiques de l’oxygène. Citons comme enzymes : la superoxyde dismutase (SOD), la peroxydase, la catalase. Cependant ces enzymes peuvent elles-mêmes être dégradées par ces radicaux libres, et surtout elles diminuent d’activité au cours de notre vieillissement. Les actions délétères des radicaux libres de l’oxygène appelées stress oxydant vont alors entraîner de nombreuses pathologies souvent mortelles.
Des recherches en nutrition et en pharmacologie ont proposé d’enrichir notre alimentation par des anti-oxydants qui neutraliseraient les radicaux libres
Citons les acides gras essentiels Oméga 3 et Oméga 6, les vitamines A, C, E.
En conclusion l’oxygène nous fait vivre, mais par ses dérivés en radicaux libres nous conduit à la mort.
Un jour les progrès de la médecine permettront d’éliminer de notre organisme la production de radicaux libres de l’oxygène.
Mais l’homme doit absolument préserver les organismes marins et terrestres contenant de la chlorophylle qui sont les poumons de notre planète. Ces poumons sont les forêts anciennes type forêt amazonienne et surtout le picoplancton marin constitué de cyanobactéries.
Dans le cas de leur disparition par la pollution engendrée par l’homme, la vie de tous les êtres consommant l’oxygène disparaîtra et comme au commencement, seules les bactéries anaérobies subsisteront.
Robert Engler. Association Mycologique et botanique de l’Hérault et des Hauts Cantons. Bulletin N° 17. 2012
Une brève [et catastrophique] histoire du vert.
Le vert des végétaux, dû à la chlorophylle, permet la photosynthèse qui, à partir du gaz carbonique atmosphérique et de l’énergie lumineuse, fabrique des molécules carbonées, bases du vivant, et des molécules riches en énergie. Je propose, au prix de nécessaires simplifications, de conter l’histoire des organismes chlorophylliens et de leur influence sur les climats. […]
La photosynthèse est apparue il y a quatre milliards d’années chez des algues bleues, en réalité des bactéries (Cyanobactéries) d’après les nouvelles classifications, et sans doute à peu près en même temps chez divers organismes unicellulaires, dans une ambiance étouffante due au gigantesque effet de serre causé par l’énorme concentration en gaz carbonique (dioxyde de carbone, CO2), une teneur de 30 % soit mille fois l’actuelle.
La source de dioxyde de carbone est le dégagement volcanique, qui n’a cessé de faiblir au fur et à mesure des temps géologiques.
Le taux d’humidité élevé, les nuages épais, et le taux d’oxygène infinitésimal qui régnaient alors ont sans doute dicté les caractéristiques de la photosynthèse.
Commençons par une caractéristique sans conséquence néfaste. Le rendement maximum de la photosynthèse intervient dès le premier pour cent de l’énergie lumineuse atteint, une nécessité sous le couvert nuageux hyperdense d’alors, qui plus est dans l’eau de mer, laquelle filtre une part importante de l’énergie lumineuse dès que la profondeur s’accroît.
Ainsi, en théorie, une plante de sous-bois ne devrait pas rencontrer de difficultés de croissance puisqu’arrive au sol environ 1 % de l’énergie lumineuse. Mais deux raisons viennent s’y opposer. Tout d’abord, l’essentiel des radiations utiles à la photosynthèse, bleu et rouge, a déjà été utilisé. Ensuite, la disposition des feuilles n’est jamais optimale, les deux tiers de l’énergie lumineuse étant perdues. En pratique, il subsiste à peine 1/3 de l’énergie solaire, qui plus est à 80 % dans le vert, qui n’est pas absorbé (c’est pourquoi les feuilles nous paraissent vertes).
La suite est moins rose. Tout d’abord, la photosynthèse, née dans l’eau, en consomme beaucoup, de par sa spécificité biochimique, puisque la photosynthèse est une oxydation de l’eau, et de par la diffusion du CO2, également dans l’eau, et qui aujourd’hui encore nécessite de l’eau.
La réaction chimique se résume ainsi : l’eau est oxydée avec l’aide de l’énergie lumineuse, oxydation qui libère des électrons lesquels, grâce à une chaîne de transferts d’électrons, permettent de former des composés riches en énergie. Ces composés sont par la suite dégradés et l’énergie ainsi libérée permet de carboxyler le CO2, c’est-à-dire de le transformer en matière organique, sous la forme de sucres à 3 carbones, chez les plantes dites C3. Mais le CO2 doit donc parvenir au bon endroit et être utilisé de suite.
La diffusion du CO2 ne comporte aucune difficulté pour un organisme unicellulaire en milieu aqueux. Il n’en va pas de même pour les plantes, qui absorbent le CO2 par des stomates sous les feuilles. Les stomates ne sont pas de simples ouvertures car leurs sont associées des chambres stomatiques saturées en vapeur d’eau. Le CO2, qui se dissout d’abord dans la vapeur, passe ensuite dans le liquide intracellulaire.
Ainsi, autant la photosynthèse est adaptée au milieu aquatique, autant elle ne l’est pas à la vie terrestre. Hélas, aucun nouvelle voie ne s’est développée chez les végétaux, à part deux améliorations.
La voie dite des plantes C4 disjoint l’oxydation de l’eau de la carboxylation (assimilation du CO2) et permet ainsi d’utiliser le CO2 plus profondément dans les tissus, et donc d’en stocker dans davantage de cellules. La photosynthèse peut alors se poursuivre en milieu de journée par temps chaud, stomates fermés. Les plantes C4 (Maïs, Sorgho) continuent de pousser en plein midi quand les autres, les C3, la grand majorité, se mettent à l’arrêt à partir de 30 °C. Au soleil, une telle température est atteinte chez nous l’été de 10 h à 18 heures et nos légumes ne poussent guère…
Autre amélioration, celle des plantes dites CAM, qui concerne des plantes crassulescentes, chez lesquelles la disjonction entre l’oxydation de l’eau et la carboxylation est temporelle. Le CO2 est alors assimilé la nuit quand les stomates peuvent rester grand ouverts.
Ensuite, la photosynthèse se caractérise par l’émission d’oxygène.
Or il s’agit d’un corps chimique hautement toxique, du fait de son pouvoir oxydant élevé, au point qu’il a donné son nom au processus d’oxydation. Songez qu’il corrompt le fer, qui rouille, et qu’il détruit nos cellules par l’intérieur, lors du processus de vieillissement, d’où la vente à prix d’or d’anti-oxydants par des commerçants avisés. Ajoutons que les plantes actuelles consomment une énergie non négligeable à éliminer l’oxygène des cellules. Il est donc probable que dans une atmosphère oxygénée, une autre forme de photosynthèse aurait été sélectionnée ; mais, dans l’ambiance quasi anoxique originelle, la toxicité de l’oxygène s’avérait sans grand risque, en tout cas sans pression de sélection.
En outre, les formes de vie demeurant à l’époque limitées, avec à la fois peu d’individus et peu d’espèces, le tout dans une abondance de ressources, aucune compétition n’existait, donc aucune pression de sélection autre que celle exercée par le milieu physique et chimique.
Enfin, la photosynthèse consomme beaucoup de CO2, un point certes inévitable puisqu’il est transformé en matière organique mais qui, en l’absence à l’époque de capacités de recyclage (chaînes de décomposition) s’est traduit par de grandes pertes de carbone, dont nous brûlons une partie aujourd’hui, et dont une partie a été dissoute dans les océans (coquilles d’organismes morts) ou stockée au fond (voir le problème des clathrates en fin d’article).
Les volcans constituent la source unique de CO2, tout le reste du cycle du carbone consiste en une consommation et un recyclage partiel, soit directement par les végétaux et le vivant (atmosphère, sols, etc.), soit, pour sa plus grande part, via les océans et les roches sédimentaires sous forme principalement de carbonates, qui trouvent leur origine dans le vivant ou son action. Si on représente le cycle du carbone de manière statique, comme dans TOUS les schémas proposés sur Internet ou dans les livres, les végétaux et le vivant apparaissent peu ; par contre, si on tient compte de l’histoire du cycle, les végétaux représentent alors le flux majoritaire, un point oublié du fait de la représentation statique – comme quoi, un simple schéma peut influencer la manière de penser !
Dans une autre atmosphère, une pression de sélection aurait sans doute abouti à un processus moins dispendieux, gaspillage éhonté, mais avec une concentration atmosphérique de 30 %, nul besoin d’économiser le CO2 n’est intervenu.
Il est d’ailleurs à noter que d’autres voies chimiques existent, chez des bactéries, aussi bien quant à la fourniture d’énergie que dans la manière de créer de la manière organique ; mais ces voies, moins efficaces et limitées quant à leurs ressources (H2S, carbonates dissous), sont demeurées minoritaires.
Par ailleurs, la vie demande divers minéraux, dont notamment l’azote (sous forme d’ammoniac ou de nitrate selon le cas), abondant dans l’atmosphère mais indisponible dans les roches (présent à l’état de traces dans les roches magmatiques, il n’existe que dans les roches sédimentaires sous des formes inatteignables à la vie). Face à un tel besoin (pas de protéines sans azote), par chance et par nécessité, est apparue presque en même temps que la photosynthèse la possibilité d’assimiler l’azote atmosphérique. La réaction chimique, liée aux conditions à cette époque, demande l’absence d’oxygène (anaérobie stricte), condition qui deviendra un problème par la suite.
Seules les Légumineuses et environ deux cents espèces, de diverses autres familles, dont l’Aulne et l’Argousier, ont trouvé la parade, par la mise sous séquestre des bactéries dans des nodules en partie étanches, qui plus est, chez les seules Légumineuses, nettoyées de leur oxygène par une substance proche de l’hémoglobine, la leghémoglobine.
Quoi qu’il en soit, malgré les défauts de la voie photosynthétique, la vie s’est développée.
Le processus semble même miraculeux, d’où notre manque de recul habituel à son propos. Pensez-donc, n’est-il pas magique que des organismes assimilent l’énergie lumineuse, surabondante, et un gaz, surabondant, et ainsi, d’amour et d’eau fraîche, en quelque sorte, créent la matière organique, base du vivant ? Un tel procédé, quasiment divin, fait oublier ses inconvénients, de même que la nécessité de l’azote pourtant peu abondant, un talon d’Achille donc. Mais les plantes et la photosynthèse continuent aujourd’hui, même et surtout en cours de sciences naturelles, à être présentées comme des miracles, l’exemple même de la nature bienfaisante et autres fadaises rousseauistes.
Pendant longtemps, la concentration atmosphérique de CO2 a légèrement diminué tandis que celle de l’oxygène augmentait, des changements presque imperceptibles vu la faible biomasse des organismes impliqués. Au bout de 1,5 milliards d’années toutefois, le dégagement d’oxygène a été suffisant pour éliminer par oxydation le méthane, puissant gaz à effet de serre (dix à vingt fois plus que le CO2), d’où une diminution dudit effet, accentuée par la diminution de la teneur en CO2, situation qui, additionnée à des causes volcaniques, a produit la première glaciation, la plus terrible d’ailleurs, il y a 2,5 milliards d’années, la glaciation huronienne.
La planète a été entièrement recouverte de glace sans le moindre endroit d’eau ou de roche libre pendant trois cent millions d’années. Il semble que la perte d’effet de serre due à l’oxydation du méthane ait été accrue par une plus faible activité volcanique, avec donc moins de libération de CO2.
Des éruptions volcaniques ayant rechargé l’atmosphère en CO2, les chlorophylliens ont repris leur travail dès la glaciation terminée, et le dégagement d’oxygène a favorisé le développement de la vie, avec les premiers êtres multicellulaires.
Si l’oxygène est toxique (dans sa forme ionisée), il recèle beaucoup d’énergie, libérée puis stockée dans le cadre de la respiration. Toutefois, d’autres types de respiration existent, sans oxygène. Ainsi, la respiration que nous connaissons peut être vue comme l’exploitation d’une ressource abondante, ou bien comme une manière de consommer le surplus d’un toxique ; et en effet, sans respiration, les végétaux se seraient rapidement intoxiqués à l’oxygène, et nous n’aurions jamais connu de forêts ni de légumes (et pour cause puisque nous n’aurions pas même existé !).
Il y a 700 millions d’années, une nouvelle glaciation intervient, dite Varanger, dont les causes semblent être plutôt géologiques cette fois (suite à des ouvertures océaniques, précipitation de carbonates entraînant la chute de la teneur en CO2), encore que la vie ait probablement joué un rôle plus important que généralement accepté.
Puis, au Cambrien, il y a 540 millions d’années, toutes les conditions furent réunies pour un développement intense des divers organismes. L’explosion de nouvelles formes de vie, presque toutes consommatrices d’oxygène et productrices de CO2, a masqué le problème que pose la photosynthèse.
En conséquence, sans réel besoin et donc sans pression de sélection, aucune modification n’est apparue dans les mécanismes d’assimilation de l’énergie solaire (photosynthèse) et de l’azote atmosphérique (assimilation bactérienne). Peu à peu, la vie sort de l’eau (au Silurien, vers 430 millions d’années) et se développe à la surface terrestre.
La vie résout alors deux problèmes majeurs : la lutte contre la dessication (hors de l’eau… il y a moins d’eau) et contre les UV. Une épaisseur infime d’eau (un simple film) suffisait à protéger des radiations et donc des mutations, sur terre d’autres moyens doivent être inventés, pigments (flavonoïdes) et cuticules chez les végétaux, écailles, plumes ou peau couverte de poils chez les animaux ou pigmentée (mélanine) chez l’homme, ainsi que diverses stratégies d’évitement, en particulier une vie nocturne plus intense que la vie diurne.
D’une manière générale, dans tout écosystème l’essentiel de la biomasse provient des végétaux. Aussi, l’énorme développement de la vie a conduit à deux conséquences, la diminution du taux de CO2 atmosphérique et l’augmentation de la teneur en oxygène, conséquences toutes deux négatives, non seulement pour le climat mais pour les organismes eux-mêmes, à des niveaux différents.
Premier niveau, l’impact de la teneur en dioxyde de carbone, à partir d’un certain seuil, limite la photosynthèse.
Second niveau, l’augmentation de la teneur en oxygène crée la photo-oxydation (dénommée à tort photorespiration), un moyen pour le végétal d’éliminer l’oxygène en excès, mais un moyen qui consomme en pure perte une partie de l’énergie assimilée (jusqu’à un tiers).
Troisième niveau, l’abondance de l’oxygène dans l’eau et dans les sols empêche l’assimilation de l’azote (puisque le processus est anaérobie). Ainsi, l’oxygène produit par les végétaux orchestre le futur manque vital en azote qui limite leur développement.
Bien entendu, de telles limites n’interviennent au début qu’à la marge, et n’empêchent pas le développement de la vie, c’est sans doute la raison pour laquelle elles sont ignorées ou présentées comme secondaires dans l’enseignement, et dans la plupart des ouvrages.
Au Carbonifère, il y a environ 320 millions d’années, l’explosion de la vie a consommé une part importante de l’oxygène fourni par les végétaux, mais pas au point d’empêcher l’augmentation de sa teneur atmosphérique. Parallèlement, le taux de dioxyde de carbone a diminué, jusqu’à 10 % lors de la pullulation arborée carbonifère. Les végétaux étaient partis pour consommer tout le CO2 et donc se détruire eux-mêmes.
Quand on invoque l’absorption du CO2 par les végétaux, mis à part la pullulation carbonifère qui voit l’explosion de formes arborées, on songe surtout au phytoplancton, aux prairies et aux pelouses, ou encore à la toundra et la taïga qui, soit de par leur dynamique consomment beaucoup de CO2 pour les premières, soit, pour les deux dernières, séquestrent le carbone par accumulation à cause de la faiblesse de la décomposition, limitée par le froid. Il en va de même, mais dans une moindre mesure, par accumulation dans les marécages ou les tourbières, cette fois par manque d’oxygène.
Les forêts, quant à elles, consomment peu de CO2 pour les formations tempérées et n’en consomment pas voire en dégagent dans les régions chaudes (parce que la décomposition y est très rapide).
Après diverses extinctions, dues à d’autres causes, et qui ont évité finalement le problème majeur d’une glaciation généralisée, la nouveauté notable pour l’histoire de la planète intervient à la fin du Jurassique, il y a environ 160 millions d’années : l’apparition des plantes que nous connaissons aujourd’hui, les plantes à fleurs ou Angiospermes. Une chance. Pourquoi, puisqu’elles ont continué à consommer le gaz carbonique ? Eh bien parce que les plantes à fleurs devraient, du point de vue des grands phénomènes écologiques, être plutôt nommées plantes à feuilles. Car une feuille se décompose.
Revenons un peu en arrière. Pendant le Carbonifère, le carbone assimilé par les végétaux s’est accumulé, à la fois à cause des conditions anoxiques dans les vases et les tourbières, et à cause du caractère coriace desdits végétaux (les pins que nous connaissons, avec leurs dures aiguilles, en fournissent une image édulcorée). Le carbone s’accumulait, donc il ne revenait pas à l’atmosphère, qui s’en appauvrissait.
Au contraire, 170 millions d’années plus tard, les feuilles tendres des Angiospermes forment des litières que des myriades de décomposeurs transforment, libérant du CO2 dans l’atmosphère ; et, au passage, créant les sols que nous connaissons aujourd’hui, dont nous héritons, et qui nous ont permis quelques milliers d’années d’agriculture facile, jusqu’à leur épuisement en un petit siècle avec la révolution agricole.
Cependant, les Mésopotamiens avaient déjà transformé leur Croissant fertile en désert par des irrigations intempestives qui ont causé une salinisation en surface, et les Africains avaient créé le Sahel à partir d’une quasi forêt, qu’ils ont méthodiquement brûlée. Les Européens n’ont dû la relative pérennité de leurs terres qu’à leur climat tempéré. Quant aux Indiens, ils ont réussi à stériliser plus de la moitié des terres très fertiles de la vallée du Gange. Les sols américains, eux, ou ce qu’il en reste, sont sous perfusion. Les plantes à feuilles constituent la meilleure, et pour tout dire la seule, invention des végétaux propre à limiter les pertes de CO2, en le recyclant dans des chaînes de décomposeurs.
Sans eux et sans les feuilles, la terre ne serait plus qu’une boule de glace depuis au moins cent millions d’années.
Bénissons donc les sols, et leur cortège de bactéries, champignons, protozoaires, nématodes, insectes, vers… Mais, si nous tenons à limiter l’effet de serre, une attitude absurde comme nous le verrons, nous pourrions utiliser un moyen idiot mais simple, peu coûteux et radical : anéantir la vie des sols. Curieux que personne n’y ait encore songé…
Bon an mal an, le taux de CO2 atmosphérique a donc diminué moins vite que l’assimilation photosynthétique ne l’aurait fait craindre, mais il a diminué tout de même jusqu’à une valeur que nous connaissions avant la révolution industrielle, moins de trois pour dix mille (0,027 pour cent ou 270 ppm).
L’action de l’homme a porté la concentration à 410 ppm soit, à l’échelle du vivant, une modification infime, bien qu’assez pour modifier légèrement le climat.
Autant dire que, par rapport aux 30 % de départ, la teneur atmosphérique récente en CO2 en fait un gaz résiduel. Le CO2 est devenu alors LE facteur limitant du développement de la vie végétale, et donc par ricochet de la vie animale.
On pourrait juger osé à première vue de ramener le cycle du carbone au seul vivant.
Mais d’une part les entrées par les volcans sont faibles sur un temps court, d’autre part, si d’autres causes interviennent dans les bilans, en particulier la dissolution ou non du CO2 dans l’eau avec éventuelle séquestration sous forme de carbonates, ces causes connues et prévisibles n’expliquent pas les variations… sauf quand elles sont dues à l’activité du plancton (par exemple pour la précipitation de la craie).
Parallèlement, du fait de la haute concentration d’oxygène dans l’atmosphère (21 %), l’assimilation d’azote atmosphérique devenait anecdotique, une carence heureusement atténuée, mais pas compensée, par la décomposition de la matière organique permise, donc, par la nature tendre des feuilles (le recyclage animal compte pour 10 % dans le bilan), ainsi que par le développement des Légumineuses.
Sans doute étions-nous arrivés au taux minimum de CO2 compatible avec le couvert végétal de la planète.
On estime d’ailleurs que la teneur actuelle en CO2 atmosphérique divise par cent le potentiel de croissance des végétaux, mais d’autres causes interviennent également, comme le manque d’azote.
Nous sommes alors au Quaternaire où, justement, le fait de parvenir à une telle limite va déterminer les climats : les glaciations et âges interglaciaires se succèdent à des vitesses incompatibles avec les temps géologiques, mais en harmonie avec l’évolution des forêts, quelques centaines d’années ; toutefois, dans l’hémisphère Nord seulement. Pourquoi ?
Outre diverses autres raisons (en écologie, les causes s’avèrent toujours multiples et intriquées), les végétations (forêts, mais pas seulement) au Nord et au Sud fonctionnent tout à fait différemment. Au Nord, du fait de températures peu élevées, la décomposition lente crée des sols mais y séquestre le carbone, d’où une diminution de l’effet de serre, suffisante pour enclencher une glaciation. Au Sud, du fait des températures élevées, la décomposition est si rapide qu’aucun sol ne se crée. Le carbone n’est pas séquestré, l’effet de serre perdure. On peut ajouter que la plus grande humidité atmosphérique des forêts équatoriales, dites ombrophiles, ou forêts humides, contribue également à l’effet de serre (l’humidité à saturation est due à la densité du couvert végétal qui empêche toute circulation d’air).
Le mécanisme, bien que mal connu dans les détails et insuffisamment documenté dans ses déterminants, est donc le suivant : une fois parvenu à un quasi épuisement du CO2, l’effet de serre disparaît, la température chute, la végétation meurt. L’augmentation de l’albedo qui en résulte (une forêt ou une prairie reflètent moins la lumière solaire qu’un sol nu ou, pire, enneigé) accroît la chute de température. Une glaciation se produit, le CO2, qui n’est plus consommé, mais continue à être restitué à l’atmosphère par divers mécanismes, décompositions, océans, éruptions volcaniques, accroît peu à peu sa teneur ; l’effet de serre augmente, la température également, l’interglaciaire arrive, devient vite chaud. Une explosion de la vie se produit, principalement chlorophyllienne puisque la biomasse végétale représente 90 % du total ; par suite, le taux de CO2 baisse et enclenche la future glaciation.
Certes, le Quaternaire correspond par ailleurs à un minimum de température sur le plan géologique, minimum mal expliqué mais probablement dû à des causes cosmotelluriques.
C’est justement l’existence d’un tel minimum qui rend critique le niveau de CO2 atteint en grande partie sous l’influence des végétaux. Dans une période géologique chaude, le problème serait minimisé voire masqué. Si une telle période intervient, l’évolution se fera sur des temps géologiques, c’est-à-dire sur des millions et plutôt des dizaines ou centaines de millions d’années. Ainsi, l’impact géologique (cosmotellurique) peut être supérieur à l’effet végétal ou le déterminer, mais sur des échelles de temps sans aucune commune mesure.
À l’époque actuelle, nous étions toujours bloqués à l’intérieur d’un tel mode oscillatoire, à la fois par la pénurie en CO2 et par celle en azote due à l’excès d’oxygène. On peut dire que les végétaux ont modifié la planète au point d’en parvenir à se limiter eux-mêmes. À conditions géologiques égales, rien n’aurait empêché les cycles glaciaires de s’enchaîner indéfiniment. Mais l’homme a consommé en un siècle une petite part du carbone stocké au carbonifère, rejetant le CO2 et augmentant l’effet de serre. L’augmentation de la teneur en CO2 est très favorable à la vie. L’effet de serre perdurera car les végétaux ne consommeront pas tout, de plus un effet retard existe, du fait que nous étions à une époque interglaciaire avec une couverture végétale importante, du fait également que l’azote manque, et enfin que la teneur en CO2 demeure modeste, mille fois moins qu’à l’origine. Rien ne pouvait arriver de mieux à la vie qu’un relarguage intensif de CO2 dans l’atmosphère…
On peut en conclure que les plantes ont modifié, voire détruit, la planète. On constate le même phénomène avec tous les organismes importants, bactéries, champignons, plancton, qui modifient le milieu à leur avantage, mais au détriment des autres formes de vie, et finalement à leur propre détriment. L’homme ne constitue donc pas une exception, constat qui rend pessimiste quant à sa capacité à se sortir du piège dans lequel il s’est fourré : en toute logique, il ira jusqu’à la destruction totale de ses conditions de vie.
Ajoutons pour noircir le tableau que tout organisme épuise les ressources dont il dispose s’il n’est pas limité par des prédateurs, des parasites ou d’autres causes. De simples bactéries, si elles n’étaient pas dévorées avant, consommeraient en moins de 24 heures l’ensemble du vivant de la planète… Il nous aura fallu deux siècles !
Bien sûr, on peut également voir les choses autrement : les végétaux ont créé tous les milieux de vie qui existent à la surface de la planète et, sans eux, la diversité serait probablement moindre, voire minime. Tout dépend en fait de l’échelle à laquelle on raisonne : à l’échelle de la planète, les végétaux ont épuisé leurs ressources ; à l’échelle d’une forêt, d’une prairie ou d’un sol, ils ont créé et entretiennent un milieu de vie. Qu’on détruise la forêt et le sol s’érode rapidement, l’essentiel de la vie est alors anéantie. Aussi n’y a-t-il pas incompatibilité entre les deux raisonnements, l’un qui voit (à grande échelle) les végétaux comme responsables de cataclysmes planétaires, l’autre (à petite échelle) qui les considère comme la matrice de la vie.
Note : jusque récemment, les causes invoquées des glaciations étaient multiples, mais aucune explication ne satisfaisait personne. Les glaciations apparurent dans des configurations cosmiques et géologiques diverses, qui les éliminent donc en tant que facteurs déterminants. Elles coïncident par contre avec les fluctuations des teneurs atmosphériques en CO2 et en O2. On alors tenté d’expliquer lesdites fluctuations par des causes cosmiques ou géologiques, sans succès. La seule cause majeure des changements des teneurs en CO2 et en O2 est le vivant. Rappelons aux sceptiques que malgré le quasi minimum que représente l’effet de serre actuel, sans lui, la température moyenne du globe, indépendamment de la latitude et de la saison, serait de – 18 °C. Ainsi la photosynthèse, à elle seule, détermine la température moyenne et donc le régime glaciaire ou non. Les phénomènes cosmiques et géologiques accroissent ou limitent les effets mais ne les déterminent pas. Par contre la géologie, notamment par la position des continents, détermine les climats locaux (local à l’échelle de la planète, donc au sens de continental).
Bien sûr, l’effet de serre nous sera moins favorable, à nous, humains. Espérons toutefois que l’humanité, si elle survit, n’arrêtera pas totalement de dégager du CO2 dans l’atmosphère, au risque de créer une glaciation dans quelques centaines d’années… Ce serait un comble, non ?
L’action de l’homme, elle n’a d’effet que sur lui-même (du moins si on la limite au dégagement de CO2 et qu’on met temporairement de côté la chimie et les destructions), et encore l’effet n’est-il que géopolitique (gérer des déplacements de populations). Mais, si les choses étaient faciles, nous pourrions tous migrer, en quelques générations donc de manière indolore, vers les toundras actuelles : l’exploitation des terres libérées par le dégel du permafrost nous offrira une agriculture prospère et sans nécessité d’intrants pendant au moins mille ans (et plutôt dix mille, même en les surexploitant à outrance).
Malheureusement, la propriété privée et les revendications des états compliqueront les choses, d’autant que l’immense majorité desdites terres se situe en Russie……
Le permafrost (ou pergélisol) résulte de l’accumulation de matière organique peu décomposée à cause du froid qui ralentit l’activité microbienne, puis de sa prise en masse par le gel sur de grandes épaisseurs, jusqu’à 500 mètres. Le méthane, qui résulte de la dégradation anaérobie de la matière organique avant le gel, conditions anaérobies dues à l’accumulation de matière organique, s’organise avec la matière organique sous forme de clathrates, stables. Un tel piégeage de la matière organique existe également dans les fonds marins.
Chaque m³ de clathrates renferme 0,164 m³ de méthane, gaz à effet de serre 10 à 20 fois plus puissant que le CO², rappelons-le. Si le permafrost fond, il sera rapidement exploitable car le dégagement de méthane créera un emballement de l’effet de serre et une augmentation très rapide de la température, de l’ordre de 5 à 15 °C en moins d’un siècle, augmentation extrêmement favorable à la vie (en moyenne, mais pas partout ni toujours). Ainsi sera le monde si une partie du CO2 séquestré par l’intermédiaire de la photosynthèse retourne à l’atmosphère sous sa forme initiale ou via le méthane.
L’homme n’est pas le seul à disposer du pouvoir de causer la libération des clathrates.
Dans un passé récent, un volcan sous-marin a fondu des clathrates avec à la clé un réchauffement rapide de 5 °C (Maximum thermique du Paléocène tardif il y a 55 millions d’années). Nous sommes donc assis sur des bombes à retardement que nous laissent les végétaux à force d’avoir dilapidé le CO², désormais stocké un peu partout sous forme de matière organique…
Didier Vereeck, Association Mycologique et botanique de l’Hérault et des Hauts Cantons. Bulletin N° 22. Mai 2017
Les grands changements climatiques vont être dus pour le principal à l’excentricité de l’orbite terrestre : variation de l’axe des pôles par rapport au plan de l’écliptique – le plan sur lequel la terre tourne autour du soleil – . On parle aussi de variations de la circulation des courants océaniques : la convection thermohaline.
Sur le site actuel d’Oklo, proche de Franceville, au Gabon, se mettent en activité en 16 endroits différents des réactions de fission nucléaire en chaîne auto-entretenue dans une veine d’uranium encadrée de grès et de granits. C’est un phénomène naturel, du au ruissellement des eaux de surface sur l’uranium, unique à notre connaissance, tout à fait identique à ce qui se passe dans nos réacteurs nucléaires, en beaucoup moins puissant, qui sera découvert en juin 1972, et qui aura cessé quand commencera l’exploitation de l’uranium au XX° siècle. C’est à dire que cette pile naturelle aura produit pendant deux milliards d’années des déchets nucléaires qui n’ont pas perturbé outre-mesure les vie locale, végétale, puis animale et enfin humaine.
L’idée d’une espèce de réacteur nucléaire (d’origine non humaine) fonctionnant à l’intérieur de la Terre peut sembler absurde. Pourtant, elle ne l’est pas… Il existe en effet une preuve indiscutable que le phénomène est possible. Dans la mine d’uranium d’Oklo, en République Gabonaise, ont été retrouvés les résidus d’une réaction de fission nucléaire auto-entretenue s’étant déroulée dans la roche il y a environ 1,7 milliard d’années.
La découverte date de juin 1972, lorsque l’on a constaté qu’une partie du minerai était anormalement pauvre en 235U. Il fallut rapidement se rendre à l’évidence, comme le montrèrent les analyses ultérieures : pendant plus de 400.000 ans une série de petits réacteurs nucléaires naturels avaient spontanément démarré puis fonctionné pendant de longues périodes. Originellement dispersé dans du grès, de l’uranium 235 s’était retrouvé rassemblé dans certaines zones jusqu’à atteindre la concentration nécessaire au démarrage de la fission. La présence de circulation d’eau avait servi de modérateur et permis aux réactions en chaîne de se produire lentement mais de façon stable.
https://matierevolution.fr/spip.php?article4338. 1 février 2017
ère géologique : Stathérien : 1.8 à 1.6 milliard d’années
1.75 milliard
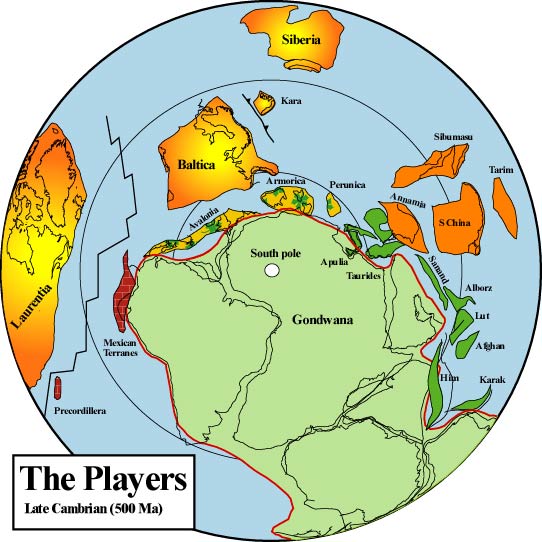
Première fragmentation de la terre émergée, qui donne naissance à deux ensembles continentaux : la Laurasie (aujourd’hui Amérique du Nord, Europe du sud, Asie) et le Gondwana (aujourd’hui Amérique du Sud, Afrique, Inde/Madagascar, Arabie, Australie, Antarctique), qui vont être séparés par un océan, zone de fractures : le Paléo-Téthys. On voit circuler plusieurs noms sur ces premiers âges de fragmentation des terres ; ils correspondent en fait à des périodes différentes, les unités de temps utilisées étant plutôt de l’ordre du milliard d’années que du siècle : il faut surtout garder à l’esprit que, dès cette première fragmentation, la croûte terrestre n’a jamais cessé de se fractionner, tel un puzzle qui se déferait et se referait indéfiniment ; ainsi, il y aurait eu en premier lieu un continent unique nommé Nuna – ou Hudsonie -, de 1,8 à 1,5 milliard d’années, suivi d’un autre, Rodinia, de 1,5 milliard d’années à 750 m.a., lequel engendrera la Pangée qui engendrera le Proto Laurasie, soit en gros, l’hémisphère nord et le Proto Gondwana, – l’hémisphère sud -, ce dernier devenant le Gondwana vers 510 m.a. qui se fracturera au début du Crétacé, vers 136 m.a. Ces assemblages de cratons ne se font pas comme celui d’un puzzle dont les pièces ont été prédécoupées pour s’ajuster avec précision les unes aux autres, mais dans une grande bousculade de plaques qui se rencontrent avec une puissance inimaginable, s’enchevêtrant les unes les autres ; pour le Gondwana, c’est l’origine des montagnes du cap Horn pour l’Amérique du sud, du Cap pour l’Afrique du Sud, des îles Shetland pour l’Antarctique.

Cratons du Mésoprotérozoïque (+ de 1,3 Ga) en Afrique et Amérique du Sud.
http://www.syti.net/EvolutionStory.html
1.7 milliard
Les roches sédimentaires qui se sont déposés à l’emplacement de l’actuel cañon du Colorado se métamorphisent sous l’action de la chaleur du magma et de la pression. Ce sont les schistes et les grès que l’on retrouve aujourd’hui tout au fond du cañon.
Mésoprotérozoïque : 1,6 à 1 milliard d’années.
1.6 milliard Premières cellules eucaryotes (avec un noyau) : ancêtre commun aux animaux, aux plantes, aux champignons et aux protistes.
ère géologique : Calymnien : 1.6 à 1.4 milliard d’années.
ère géologique : Ectasien : 1.4 à 1.2 milliard d’années.
1.3 milliard Des algues productrices de chlorophylle a et de chlorophylle b donnent naissance aux premières algues vertes.
ère géologique : Sténien : 1.2 à 1 milliard d’années.
1.2 milliard Une mer peu profonde occupe l’actuel site du cañon du Colorado, dans le fond de laquelle s’accumulent 4 000 mètres de sédiments et de laves volcaniques.
La grande aventure de la vie sur Terre
5. Il y a 1.2 milliard d’années, les premières algues et la photosynthèse.
On a longtemps considéré les cyanobactéries comme des algues, peut-être les toutes premières. Les scientifiques savent aujourd’hui qu’elles en sont plutôt les précurseurs, car leur structure est celle des bactéries, ou procaryotes, c’est-à-dire des organisme unicellulaires dépourvus de noyau. Leur capacité à exploiter la lumière du soleil pour se fournir en énergie va cependant bel et bien précipiter l’apparition de tout un pan du règne du vivant : les plantes. La première fut sans nul doute une sorte d’algue unicellulaire ; un eucaryote primitif qui assimila une cyanobactérie par endosymbiose. Cette bactérie devint peu à peu la structure qu’on appelle aujourd’hui chloroplaste, l’organite permettant la photosynthèse chez les plantes et les algues.
William Rowe-Pirra. Sciences et Avenir n° 208 de janvier à mars 2022
ère géologique : Tonien : 1 milliard d’années à 720 millions d’années
1 milliard
Les conditions thermodynamiques au niveau de l’interface lithosphère/manteau terrestre qui permettent les phénomènes de subduction, sont en place.
Les roches passent d’une forme à une autre selon un processus continu, ou cycle des roches. En surface, les roches subissent l’érosion ; devenues sédiments, elles sont transportées et cimentées en une roche sédimentaire. Celle-ci peut être transformée par la pression et la chaleur en une roche métamorphique, puis fondue en magma et rejetée enfin à la surface sous forme de lave volcanique. Dès que le magma se solidifie en une roche ignée et que celle-ci est exposée à l’air et à l’érosion, le cycle recommence.
[…] Les roches ignées résultent de la solidification du magma. Expulsé par un volcan, le magma peut prendre plusieurs formes, de l’obsidienne vitreuse à la pierre ponce. Stagnant sous la croûte, il forme un pluton qui refroidit lentement en roche intrusive ou plutonique fortement cristallisée, le plus souvent un granite, composé de quartz, feldspath alcalin et plagioclase [mica].
Nouvel Atlas Universel. Reader’s Digest. 1998
Avec le basalte, le granite est la roche la plus répandue à la surface du globe : 43 % en basalte, qui tapisse plutôt le fond des océans et 22 % en granite, plutôt sur les continents. Les granites constituent les produits les plus évolués et les plus tardifs de la cristallisation des magmas silicatés. Cette cristallisation peut se former à des températures de l’ordre de 700 °C facilement atteintes dans la croûte, à condition que les teneurs en eau du milieu soient suffisantes. Certains granites apparaissent en milieu de plaque ou en zone de divergence de plaques, c’est-à-dire dans une région non soumise aux effets orogéniques : ils sont anorogéniques. Les autres, majoritaires, s’installent dans les chaînes de montagne au moment de leur formation et sont orogéniques.
Les granites participent à la croissance et au recyclage des océans. À l’échelle du globe, la production de magmas provenant du manteau est estimée à 30 km³ par an en moyenne.
Aujourd’hui, on établit trois grandes catégories de roches :
- Les roches magmatiques ou ignées ; venues d’un magma en profondeur, ce sont des roches plutoniques : granite, diorite, andésite ; venues d’une zone plus proche de la surface, ce sont des roches volcaniques : basaltes, laves.
- Les roches sédimentaire, formées sur le fond des océans par décomposition des coquillages : grès, calcaires, argiles, marnes.
- Les roches métamorphiques : préexistantes à grande profondeur et cuites à haute température : schistes, houille, serpentine.
800 m.a.
La température des océans est d’environ 20°.
750-700 m.a.
Glaciation dite de Ghaub [ou Varanger…selon le pays d’où l’on en parle] : on en trouve des traces en Namibie, alors dans l’hémisphère sud à des latitudes tropicales : les variations climatiques tenaient déjà dans ces temps lointains essentiellement aux variations de l’intensité de l’effet de serre. Les algues partent à la conquête des océans.
725 m.a.
Une chaîne de montagnes se forme à l’emplacement de l’actuel cañon du Colorado, suivie de phases d’érosion puis de submersions marines de 550 à 250 m.a.
ère géologique : Cryogénien : 720 à 635 millions d’années
700 m.a.
Le bloc australien se soude au bloc antarctique.
ère géologique : Édiacarien : 635 millions d’années à 541 millions d’années
600 m.a
Premiers métazoaires, dont les fossiles seront découverts à Ediacara, en Australie : des disques dont les plus grands atteignent un mètre de diamètre, contre seulement 6 mm d’épaisseur : ainsi l’absorption de la lumière était maximale pour assurer la photosynthèse des algues qui leur étaient liées. D’autres pluricellulaires apparaissent : vers, coraux, éponges, méduses etc…
La terre connaît un épisode glaciaire global : on en retrouve des témoignages jusqu’en Afrique intertropicale.
580 m.a
Les conditions de l’explosion précambrienne se mettent en place, mais encore aujourd’hui, plusieurs hypothèses sont émises :
-
Un réchauffement important de l’atmosphère terrestre due à l’augmentation de teneur en dioxyde de carbone, CO² et donc de l’effet de serre, et ceci en une centaine d’années, faisant fondre une grande partie de la glace et libérant ainsi des formes de vie jusqu’alors emprisonnées. La prolifération des plantes aurait alors entraîné une augmentation de la teneur en oxygène, et donc du développement du vivant.
-
La chute d’un astéroïde, Acraman en Australie, par 32°1’ S et 135°27’ E, qui se serait écrasé à une vitesse de 90 000 km/h (25 km/sec), creusant un cratère de 4 000 m de profondeur, engendrant tremblements de terre et tsunamis de plus de 100 m de haut. On a retrouvé des éjecta, grains de quartz choqués à plus de 450 km du lac salé d’Acraman, de 20 km de diamètre. Les débris arrivant à la suite auraient mis le feu au méthane des marais, provoquant des incendies dont les fumées obscurcirent le ciel pour des années, faisant chuter la température et disparaître la plupart des organismes. Seuls les survivants les plus résistants, purent recoloniser la terre, une fois revenu le retour à la normale, sans concurrence aucune de qui que ce soit.
La grande aventure de la vie sur Terre
6. Il y a 575 millions d’années, des organismes aux formes étranges
À la sortie d’une longue période de glaciation, la faune marine se développe. Si des études repoussent l’apparition des premiers animaux quelques 300 millions d’années plus tôt, une explosion de biodiversité donna alors naissance à certains des plus anciens organismes complexes connus avec (presque) certitude. Dickinsonia foliacée, Chamia à l’allure de tube ou encore Tatena inflata en forme de disque… Ils présentent des formes étranges, ce qui rend leur classification délicate. S’ils semblent avoir prospéré pendant des dizaines de millions d’années, ils furent évincés lors d’une explosion de biodiversité plus foisonnante encore.
William Rowe-Pirra Sciences et Avenir n° 208 de janvier à mars 2022
558 m.a.
Le Dickinsonia vit au fond des océans : il est plat, ovale, et n’a aucun des organes des animaux évolués mais on le dit tout de même animal, principalement parce qu’on lui aurait trouvé du cholestérol ! [si ! si ! et on ne rit pas ! Mais depuis, il prend sa dose de statines tous les jours, et ça va nettement mieux, même s’il est guetté par la maladie d’Alzheimer !]

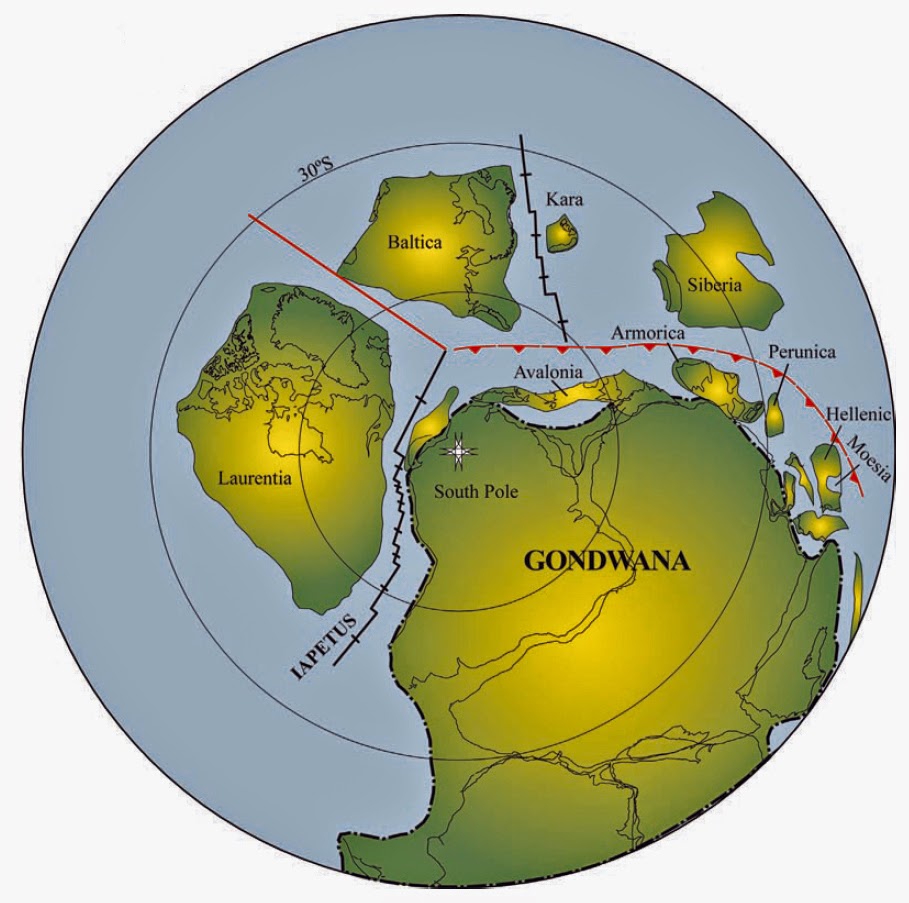
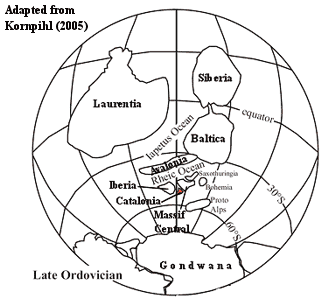
Reconstruction tardive de l’hémisphère sud au Protérozoïque Dans le dernier protérozoïque (550 Ma), Rodinia s’est fragmenté en formant Laurentia (Amérique du Nord), Baltica (Europe du Nord) et Gondwana (principalement nos continents de l’hémisphère Sud) avec l’ouverture de l’océan Iapetus. Les terranes péri-gondwanais d’Avalonia, d’Armorica et d’autres se sont rassemblés sur la marge nord du Gondwana, du craton de l’Amazonie à l’Afrique de l’Ouest. De nombreux aspects de ces reconstructions paléographiques font l’objet de débats intenses et d’investigations en cours.
PHANÉROZOIQUE. De 541 m.a. à aujourd’hui.
Le Phanérozoïque couvre la période où sont apparues des organismes pluricellulaires (métazoaires) diversifiés, pouvant atteindre de grandes tailles.
C’est à ce moment que s’est amorcée la différenciation entre les plantes et les animaux. Les premiers organismes ont alors choisi entre deux parcours de vie distincts que l’on peut, quitte à risquer un certain simplisme, résumer en ces termes : les plantes ont opté pour la sédentarité ; les animaux, pour le nomadisme. Et soit dit en passant, il n’est pas inintéressant de rappeler à ce propos que l’adoption d’un mode d’existence sédentaire a donné naissance, dans l’histoire de l’humanité, aux premières grandes civilisations.
D’emblée, les plantes se sont donc retrouvées dans l’obligation de tirer de la terre, de l’air, et du soleil, tout ce dont elles ont besoin pour vivre. De leur côté, les animaux ont été contraints de se nourrir d’autres animaux ou de plantes, et de développer des formes multiples de mouvement (course, vol, natation…). Voilà pourquoi les plantes sont dites autotrophes (du grec autos, soi-même, et trophè, nourriture) et autosuffisantes ; leur survie ne dépend pas d’autres êtres vivants. Les animaux, en revanche, sont dits hétérotrophes (du grec hétéros, l’autre, et trophè, nourriture) ; ils ne sont pas autosuffisants.
Au fil des générations, ce choix originaire a eu pour conséquences d’autres différences fondamentales entre les règnes animal et végétal, à tel point qu’aujourd’hui, on peut y voir le yin et le yang, le noir et le blanc de l’écosystème. Les plantes sont sédentaires, passives, lentes ; les animaux sont mobiles, agressifs, rapides. On pourrait multiplier à l’envi ces attributs antinomiques, et le résultat ne changerait pas ; dans le monde végétal et le monde animal, depuis cinq cent millions d’années, la vie n’a pas évolué de la même manière.
Le choix primitif entre sédentarité et nomadisme a abouti à une différentiation considérable des corps et des modes d’existence. Les animaux ont décidé de se défendre, de se nourrir et de se reproduire au moyen du mouvement (y compris, au besoin, la fuite). Les plantes ayant, quant à elles, décidé d’obtenir le même résultat en demeurant immobiles, il leur a fallu pour cela chercher des solutions tout à fait originales, du moins de notre point de vue (qui, ne l’oublions pas, reste celui d’un animal).
Immobiles et donc sujettes à la prédation des animaux, les plantes ont commencé par développer une sorte de résistance passive aux attaques venues de l’extérieur. Leur corps est ainsi formé selon une structure modulaire où chaque partie est importante sans qu’aucune soit indispensable. Cette structure offre des avantages essentiels par rapport au monde animal, surtout si l’on tient compte du nombre d’herbivores présents sur terre et de l’impossibilité substantielle d’échapper à leur voracité. Le premier de ces avantages tient ainsi à la capacité des plantes d’être mangées sans pour autant subir des dommages trop graves. Or, aucun animal ne peut en dire autant.
La physiologie des végétaux repose en effet sur d’autres principes que celle des animaux. L’évolution de ces derniers les a conduits à concentrer la quasi-totalité de leurs fonctions vitales sur quelques organes tels que le cerveau, l’estomac ou les poumons. Les plantes, au contraire, ont tenu compte de leur vulnérabilité face à leurs prédateurs, et elles ont évité de regrouper leurs facultés sur quelques espaces névralgiques. Elles se sont comportées, en quelque sorte, à la manière de quelqu’un qui, face à une forte probabilité de vol, ne rassemble pas tout son argent en un seul lieu, mais le répartit entre plusieurs cachettes, afin de limiter les pertes en cas de larcin ; ou encore à la manière des épargnants qui diversifient leurs investissements pour mieux étaler les risques encourus. […]
Chez les plantes, les fonctions ne sont pas liées aux organes. En d’autres termes, elles respirent sans poumons, se nourrissent sans bouche et sans estomac, se tiennent debout sans squelette et sont en mesure de prendre des décisions même sans cerveau.
Et c’est grâce à cette physiologie tout à fait particulière qu’un arrachage même massif ne compromet pas nécessairement leur survie : certaines d’entre elles peuvent perdre jusqu’à 90 %, voire 95 % de leur organisme et repousser ensuite, en toute normalité, à partir du petit noyau ayant survécu à la prédation. Un pâturage peut-être brouté par tout un troupeau et se régénérer en quelques jours. […] Organismes sédentaires, les plantes ont adopté, au cours de leur évolution, une stratégie qui consiste à se composer de parties divisibles pour mieux résister à leurs prédateurs. Les animaux ayant, au contraire, fondés leurs stratégies défensives en premier lieu sur le mouvement, ils n’ont développé des capacités régénératrices que dans un très petit nombre de cas. Le lézard peut certes reconstituer sa queue, lorsqu’elle est coupée, mais il ne saurait en faire autant ni de ses pattes ni de sa tête. Lorsqu’une plante subit une mutilation, non contente de survivre, elle en tire même parfois, des avantages : il suffit, à ce propos, de penser aux effets revigorants des élagages. Cette caractéristique résulte d’une structure très différente de la nôtre et constituée de modules réitérés : les branches, les troncs, les feuilles ou les racines d’une plante correspondent à autant de modules plus simples accrochés les uns aux autres tout en restant indépendantes, un peu à la manière des éléments d’un jeu de construction.
Un géranium qui orne une terrasse ne donne certes pas cette impression, et il apparaît plutôt comme un être vivant unique. Mais si vous en coupez un morceau et que vous le replantez ailleurs – si vous le bouturez, dirait un jardinier -, ce morceau détaché s’enracinera et donnera naissance à une nouvelle plante [5]. En revanche, ni un bras d’homme ni une patte d’éléphant ne peuvent régénérer un organisme entier, ou continuer à vivre séparés du reste de leur corps.
Ce n’est d’ailleurs pas un hasard si nous avons pris l’habitude de nous référer à nous-mêmes en parlant d’individus (du latin in, qui signifie non, et dividuus divisible. Notre corps, de fait, est indivisible : si l’on nous coupe en deux, nos deux moitiés seront incapables de vivre chacune de son côté et elles mourront. Si l’on coupe une plante en deux, en revanche, ses deux moitiés pourront continuer à vivre en parfaite indépendance, et ce pour une raison très simple : une plante n’est pas un individu. La manière la plus correcte de concevoir un arbre, un cactus ou un buisson, ne consiste pas à le comparer à un homme ou à un autre animal, mais de l’imaginer comme une colonie. Car il ressemble davantage à une colonie d’abeilles ou de fourmis qu’à un animal envisagé isolément.
De ce point de vue aussi, malgré l’extrême ancienneté de leur existence, les plantes témoignent d’une exceptionnelle modernité. L’un des concepts fondamentaux sur lesquels reposent les technologies rendues possibles par l’avènement d’Internet et fondées sur la connexion de groupes (par exemple les réseaux sociaux) est en effet celui des propriétés émergentes caractéristiques des super-organismes et des intelligences en essaim. On désigne par-là les propriétés que des entités individuelles ne développent qu’en vertu du fonctionnement unitaire de leur ensemble : aucun de ses composants n’est doté de manière autonome, de même justement que chez les abeilles ou les fourmis, dont les colonies développent une intelligence collective bien supérieure à celle de leurs membres pris un à un.
Stefano Mancuso, Alessandra Viola. L’intelligence des plantes. Albin Michel 2019
ERE PRIMAIRE, de 570 à 245 millions d’années. Paléozoïque
ère géologique : Cambrien 541 à 485.4 millions d’années.
La grande aventure de la vie sur Terre
7 Entre 541 et 530 millions d’années, l’explosion cambrienne
Cette nouvelle explosion va durablement marquer le monde du vivant. C’est la genèse de presque tous les embranchements majeurs d’animaux : arthropodes, mollusques, vers, éponges, échinodermes et chordés – ces organismes dotées d’une structure dorsale rigide et dont les vertébrés feront plus tard partie – y font leur apparition. Les océans fourmillent soudain d’une vie plus variée. D’une organisation simple (unicellulaires, petits organismes pluricellulaires, colonies de cellules), les êtres évoluent vers une structure beaucoup plus élaborée, avec des innovations essentielles : les intestins, la mâchoire, les yeux, ou encore un squelette interne minéralisé. Les changements sont de taille, littéralement, puisque les animaux gagnent aussi en envergure. Avec ses 80 centimètres, l’Anomalocaris était un véritable géant dans les mers cambriennes ! Mais explosion ne signifie pas chaos. L’écosystème marin, qui se complexifie conjointement, voit la mise en place d’une chaine alimentaire bien établie, enrichie de nouvelles relations de prédation.
William Rowe-Pirra. Sciences et Avenir n° 208 de janvier à mars 2022
540 m.a.
Les mers sont peu profondes et l’activité volcanique intense. Les algues vertes, sans racines, ni tiges, ni feuilles, quittent l’océan pour tenter – et réussir – la conquête des continents jusque là dénués de vie -. Ces organismes vont alors développer des racines et des tissus conducteurs de sève pour irriguer les parties aériennes. Occuper la terre ferme comportait plusieurs avantages, par exemple la possibilité de se disséminer plus facilement, puisqu’il n’existait aucune forme de concurrence. En dépit du danger de dessèchement, l’émersion permettait d’absorber plus facilement le gaz carbonique et de jouir d’un meilleur éclairage, car ces organismes avaient à leur disposition une lumière directe et non plus filtrée par l’eau. C’est ainsi que sont apparues les Bryophytes, c’est-à-dire les Mousses et les Hépatiques, toutes petites plantes primitives, encore liées au milieu aquatique, qui ne vivent actuellement que dans des milieux particulièrement humides. Des organes et des tissus se spécialisent. Les hépatiques tapissent le substrat humide pour maximiser l’absorption. Pour réduire les pertes d’eau, les épidermes sont recouverts de cutine. Les stomates – des petits trous – apparaissent pour régulariser le passage des molécules gazeuses à travers l’épiderme que la cutine a rendu étanche. Pollinisation et fécondation se font par le vent et plus tard par les insectes, mais encore – et ça, il faudra attendre 2022 pour que Myriam Valero, de la station biologique de Roscoff nous l’apprenne, en milieu marin, par l’idoté, un petit crustacé rouge de moins de 2 cm. qui assure la pollinisation des gracilaires, une algue rouge.
Apparition d’animaux complexes, à coquille rigide. On estime le nombre d’espèces animales à 160 000. Auparavant, de 3,5 milliards à 540 m.a, elles n’étaient que quelques centaines.
Le mot vie tient plus du besoin de classification de l’esprit humain qu’à une réalité physique précise : en fait, il tient à notre faculté de compréhension de la complexité des choses : tant que tout cela est à peu près compréhensible par un cerveau doté d’un QI moyen, on le nomme par le nom des principaux composants : bactéries etc… et dès que la complexification d’un organisme passe le seuil de compréhension rapide, on parle de vie.
La définition qu’en donne la NASA fait l’objet d’un telatif consensus : la vie est un système chimique capable de s’auto-entretenir et d’évoluer par sélection naturelle.
Ce qui caractérise le vivant, c’est peut-être la faculté de transmettre de l’information contenue dans l’ADN et colportée par l’ARN messager.
Didier Raoult, biologiste à Marseille.
On se retrouve donc aujourd’hui avec plusieurs scénarios, qui ne sont pas forcément incompatibles, concernant l’apparition de la vie : celui de la soupe primitive dans les océans ; celui des météorites ensemençant la Terre ; celui des sources hydrothermales dans les abysses où la pyrite de fer permet l’assemblage de molécules organiques complexes ; celui de mares de boues où l’argile, organisée en minces feuillets, facilite la production d’ARN et de petits peptides à l’abri de l’eau. Et donc celui des hydrogels d’argile, qui auraient rempli, dans les océans, cette même fonction.
Hervé Morin. Le Monde 12 novembre 2013
Et où se trouvent les frontières de la vie ? Quand on constate que des arbres sont à même de se transmettre des informations sur les prédateurs qui se nourrissent trop copieusement de leurs feuilles, ce qui leur permet d’émettre des toxines à même de détourner ces prédateurs, quand on voit la beauté ou la laideur des cristaux de glace selon l’environnement musical dans lequel ils se trouvent – voir les nombreux sites sur les expériences du Japonais Masaru Emoto -, etc… on se dit que les paramètres qui déterminent avec précision le contour du domaine de la vie demandent à être revus…
Explosion cambrienne : tous les grands embranchements animaux connus aujourd’hui sont apparus à cette occasion, apparemment sans ascendants. Les arthropodes (êtres articulés) se sont mis à proliférer : ancêtres des crustacés actuels, des araignées et des scorpions. Et aussi nombres de vers et mollusques. Tous ces animaux avaient besoin d’un taux d’oxygène plus important que les premières bactéries, car passant par l’intermédiaire de branchies ou de poumons.
À l’échelle géologique, cette explosion semble s’être déroulée en très peu de temps : tout au plus 40 m.a.
Il a fallu un coup de chance extraordinaire pour trouver les fossiles de cette époque – dans les monts Ediacara, en Australie et dans les schistes de Burgess, au Canada – : chance extraordinaire, car la plupart des fossiles sont généralement, pour les périodes les plus lointaines, des mollusques dotés d’une carapace ; or, au début du cambrien, les carapaces n’existaient pas encore. Cette faune du Burgess représente quelque chose de très spécifique dans le monde de la recherche, car elle fut décrite au début du XX° siècle comme étant bien celle de son époque et il fallut attendre les années 1970 pour réaliser qu’elle avait des caractères très novateurs par rapport aux autres faunes contemporaines. Ainsi, pour le Précambrien où la vie se limite aux bactéries, il y a 2 exceptions :
- La faune d’Ediacara constituée de métazoaires à corps mou, datée autour de 600 m.a. En 2013, des chercheurs ont trouvé dans ces schistes Spartobranchus tenuis, un ver à gland, connu sous le nom scientifique d’entéropneuste, une espèce qui prospère aujourd’hui dans le sable fin et la boue des eaux profondes et peu profondes. Selon Christofer Cameron, de l’Université de Montréal, sa description ajoute 200 millions d’années aux registres fossiles des entéropneustes, les faisant remonter jusqu’à la période cambrienne. Cela change notre compréhension de la biodiversité de cette période. Ces vers ont un corps mou, et il est extrêmement rare d’en trouver des fossiles. Les vers à gland font partie des hémichordés, avec les ptérobranches. Les Spartobranchus tenuis fossilisés sont similaires aux vers à gland d’aujourd’hui, à l’exception du fait qu’ils formaient aussi des tubes fibreux. Ils passaient ainsi au moins une partie de leur vie dans ces tubes, une habitude qui a disparu chez les vers à gland actuels, mais qui persiste chez leurs cousins les ptérobranches. Ces tubes constituent, selon les chercheurs, le chaînon manquant qui relie les deux principales familles d’hémichordés.
et, peut-être
- La faune découverte par des géologues français [ci-dessus, à 2.1 milliard] dans le Francevillien (2 000 m.a.) du Gabon s’il est confirmé qu’il s’agit bien d’organismes pluricellulaires.
505 m.a.
Premiers poissons et proto-amphibiens.
En 2014, on pêchera dans les schistes de Burgess, les fossiles d’un banc de poissons Metaspriggina jusqu’alors seulement connu grâce à deux fossiles dégradés, quand c’est une centaine de spécimens qui ont été identifiés dans les rocheuses canadiennes. Cet ancêtre putatif des vertébrés actuels est long de six centimètres, doté de branchies et d’yeux-caméras ; il devait filtrer sa nourriture sur le fond et utiliser ses muscles striés pour fuir les ancêtres des insectes et des crustacés.
500 m.a.
La grande aventure de la vie sur Terre
8 Entre 500 et 430 millions d’années, les plantes, pionnières de la vie terrestre
Les plantes seront les premières à se risquer sur la terre ferme. Grâce à une série d’innovations physiologiques, elles deviennent capables de résister aux conditions jusqu’alors meurtrières de ce milieu : l’eau y est rare, les rayons ultra-violets plus dangereux que dans les océans. Leur arsenal s’enrichit de molécules essentielles, la lignine et la cellulose entre autres. Elles permettent à ces végétaux de s’imperméabiliser et donc de réduire leurs pertes en eau, mais aussi de rigidifier leurs parties aériennes afin de pouvoir pousser à l’air libre et résister à leur propre pression sans le soutien de l’eau, ainsi que de jouir d’une meilleure protection contre les radiations venues du ciel. Les pionnières de la vie terrestre sont probablement issues d’un groupe d’algues vertes capables de survivre à des émersions temporaires grâce à un mécanisme de symbiose avec des champignons. Avec le temps, la vascularisation des plantes provoque une augmentation de leur taille et une diversification.
William Rowe-Pirra. Sciences et Avenir n° 208 de janvier à mars 2022
Les poissons, puis les amphibiens et les reptiles se dotent d’un cerveau que l’on dira archaïque qui comprend le tronc cérébral et le cervelet. Il gère les fonctions primaires liées à la physiologie de base – respiration, rythme cardiaque, pression artérielle, sommeil, équilibre et les autres fonctions physiologiques essentielles -. Seconde fonction : déclencher, face au danger, des comportements instinctifs liées à la survie, des réflexes d’attaque ou de fuite.

Je me nomme Utaurora Comosa. Je suis long de 3 cm et j’habite dans la formation Wheeler, dans le futur Utah, aux Etats-Unis.
ère géologique : Ordovicien 485.5 à 443.4 millions d’années.
480 m.a.
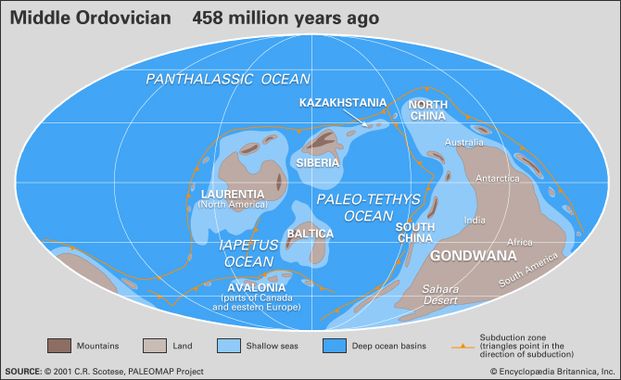
Terranne (sous-ensemble) de la Laurasie (aujourd’hui Amérique du Nord, Europe du sud, Asie) l’Avalonia s’avance en subduction sous l’Armorica vers le sud en direction du Gondwana (aujourd’hui Amérique du Sud, Afrique, Inde/Madagascar, Arabie, Australie, Antarctique), et ce dernier s’avance au contraire vers le nord ; ils sont séparés par un océan, zone de fractures : le Paléo-Téthys. Entre la couche Avalonia s’enfonçant sous Armorica et ce dernier, une zone de fusion partielle du manteau qui va créer des volcans explosifs.
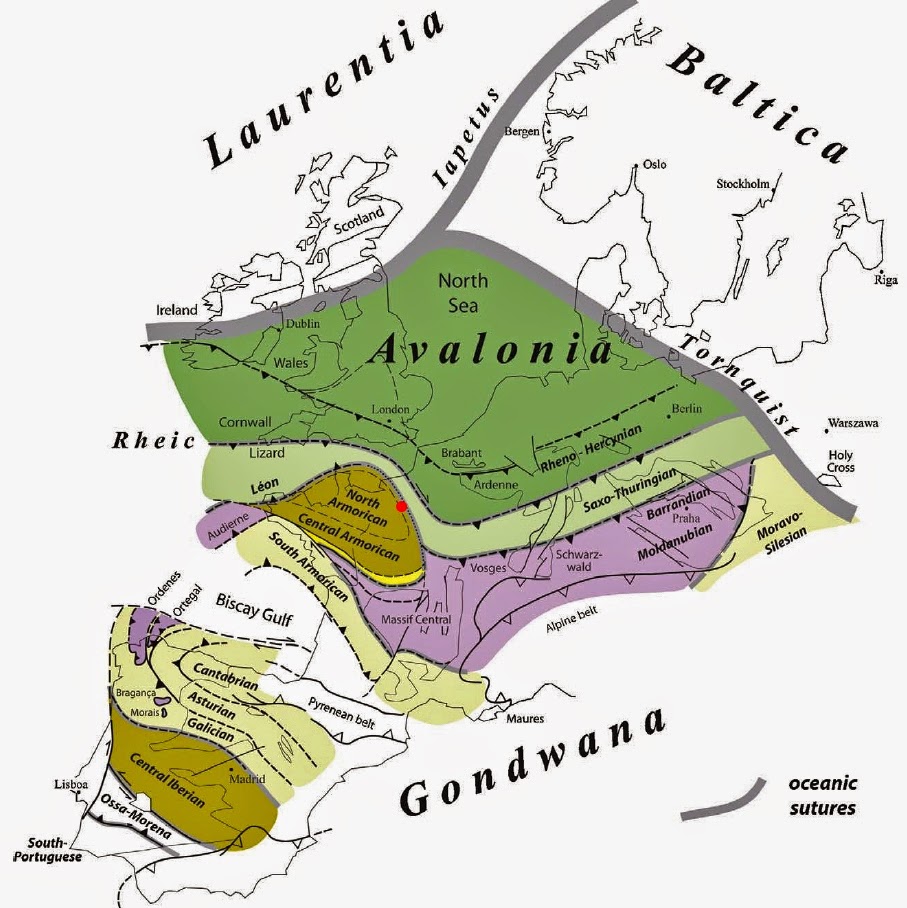
Collage européen de terranes amalgamées de type Avalonia et Cadomia, dérivées du Gondwana. Cette carte illustre l’histoire complexe de l’accrétion en Europe. Pour référence, Paris est au point rouge. Assemblage en Europe des masses continentales du Paléozoïque. La plate-forme cratonique européenne est composée d’un assemblage de terranes et de blocs de croûte continentale délimités par des failles, d’ascendance Avalonienne et Cadomienne – une Europe de croûte recyclée précambrienne et cambrienne dérivée du Gondwana. Les principaux pays qui forment l’Europe sont Avalonia, le terrane rhéno-hercynien, l’assemblage de terranes armoricains, Perunica, les Pouilles, Adria, le terrane hellénique et Moesia, tous des terranes péri-gondwanais, à l’exception de la Scandinavie dérivée de Baltica.
472 m.a.
Apparition des plantes terrestres : les premières spores fossiles sont retrouvées, en 2010, dans le nord-est de l’Argentine, alors côte nord-est du Gondwana : ces hépatiques sont de petits végétaux sans feuilles, ni tiges, ni racines, mais équipées, à l’instar des algues, d’un appareil végétatif sans cellules différenciées, ou thalle, plaqué sur le sol : c’est le premier organe de la photosynthèse. Leur ADN les relie à des petites algues d’eau douce que l’on rencontre aujourd’hui encore dans les mares temporaires, les charophytes. Des algues se seraient adaptées à l’assèchement des mares et auraient progressivement appris à vivre hors de l’eau. L’époque où elles apparaissent, l’ordovicien, coïncide avec le moment où l’oxygène s’est accumulé dans l’atmosphère pour former la couche d’ozone de haute altitude, écran indispensable à l’émergence de la vie terrestre. D’autres avaient déjà été trouvés, un peu plus jeunes, en Arabie Saoudite – 461 m.a. – et en République Tchèque – 463 m.a. -. Cinq variétés sont recensées. Il a fallu tout ce temps pour que l’évolution permette aux végétaux d’atteindre le niveau de complexité nécessaire à la colonisation des terres émergées. Ensuite, pendant à peu près 40 m.a., les plantes vont prendre de la hauteur, avec l’apparition du sporange, l’organe reproducteur de spores ; ainsi, spores et oosphères n’ont plus besoin d’eau pour se disperser : le vent peut y pouvoir ; la conquête des zones sèches peut commencer.
470 m.a.
Un fragment de météorite de 8 cm de long, baptisé Osterplana 65 et découvert en 2016 par Birger Schmitz, de l’université de Lund, vient s’incruster dans du calcaire dans ce qui est aujourd’hui la Suède à Thorsberg. Il aurait voyagé pendant un million d’années et sa datation correspondrait avec les chondrites L supposées appartenir à un astéroïde qui aurait explosé après avoir été percuté par un autre corps inconnu. Le bombardement de météorites déclenché par cette collision pourrait être responsable de l’explosion de la vie marine qui a marqué l’ordovicien.
450 m.a.
Premières plantes terrestres vasculaires, thalloïdes pour les plus simples, les autres, plus élaborées, de type Cooksonia.
445 m.a.
Des tout premiers animaux marins, combien sont parvenus jusqu’à nous, en évoluant certes, mais traversant bravement les ans par millions. Ils ne sont pas des milliers d’espèces, mais quelques unes tout de même, pour lesquelles les scientifiques ont préféré abandonner le terme fossile pour adopter celui de reliques. La plus illustre probablement : la limule.
Je pense que cela a commencé avec une pierre que j’ai trouvée dans ma maison en Pologne, où j’ai grandi. Un jour, alors que je traversais la rue en revenant de l’école, j’ai remarqué un beau coquillage parfaitement conservé, évoquant une coquille Saint Jacques, incrustée dans une grande pierre à demi-enterrée. Je n’avais qu’à la prendre et une nuit, je l’ai enterrée discrètement, une pierre énorme que je pouvais à peine soulever, et je l’ai trainée jusque chez moi. La première à remarquer ma nouvelle acquisition a été ma grand-mère. Quand je lui ai demandé ce qu’elle pensait du mystérieux coquillage, elle n’a hésité qu’une seconde avant de me répondre que c’était un vestige du Déluge, quand les animaux qui n’ont pas pu rejoindre l’arche de Noé sont morts noyés. En bon petit catholique, j’ai accepté son explication, mais en même temps, cela m’a paru paradoxal qu’un mollusque aquatique se noie dans une inondation. Je suis allé voir mon père pour clarifier la chose. En homme de science, il a sorti un livre et nous avons déterminé ensemble que la coquille appartenait vraisemblablement à un brachiopode éteint, un animal marin superficiellement semblable à un mollusque, et qu’elle devait être vieille de centaines de millions d’années. Un dessin d’une coquille remarquablement semblable à la mienne était décrite comme venant d’une période appelée Crétacé. Il y avait d’autres dessins, des libellules géantes apparemment grandes comme des oiseaux, une sorte d’immense salamandre avec la tête large et plate et des pattes courtes et trapues. J’ai trouvé cela fascinant, mais je n’arrivais pas à admettre que ces créatures merveilleuses avaient disparu et qu’on ne les reverrait jamais vivantes.
Peu après, j’ai commencé à lire des livres sur la paléontologie. La plupart étaient pleins d’informations qui passaient loin au-dessus de ma tête de huit ou neuf ans, mais les descriptions et les images de trilobites, d’ammonites, sans parler des dinosaures et des surréelles forêts de calamités, me gonflaient du désir irréalisable de remonter le temps. Puis je suis tombé sur une chose très intrigante. C’était la description d’un cœlacanthe vivant capturé au large de l’Afrique du sud – un fossile – vivant. Si ce contemporain des dinosaures avait survécu, peut-être n’était-il pas le seul. Peut-être existait-il d’autres lieux reculés – des poches de ce monde ancien – habités par des plantes et des animaux hâtivement déclarés éteints, mais toujours parmi nous.
Depuis, je suis fasciné par l’idée que certains organismes aient pu survivre plus longtemps que d’autres, et que leur apparence, et peut-être leur comportement, préserve des bribes anciennes de ce récit long et tortueux qu’est la vie sur Terre. Leur existence peut nous donner une idée des écosystèmes du passé et des interactions entre leurs habitants. Faute de machine à remonter le temps, ils sont peut-être notre meilleure chance d’appréhender les mondes fossiles disparus depuis longtemps. Beaucoup d’organismes ont reçu l’appellation excitante de fossiles vivants. Charles Darwin lui-même a été le premier à soutenir l’existence d’animaux et de plantes qui seraient restées pendant de longues périodes géologiques, conservant leur corps et faisant office d’archives du passé de la Terre.
Le concept de fossile vivant a séduit plus d’un scientifique et a nourri de nombreuses controverses dans lesquelles il est brandi comme preuve pour ou contre l’évolution de la vie.
Jeune étudiant en biologie, j’ai embrassé avec enthousiasme l’idée séduisante de créatures vivantes qui auraient résisté à la pression du changement provenant de leur environnement en perpétuelle transformation. C’étaient eux ma machine à remonter le temps, ces vestiges fossilisés rendus à la vie.
Mais en dépit de sa simplicité apparente et d’une longue liste d’animaux et de plantes qui semblent correspondre à cette définition, la validité du concept de fossile vivant n’a cessé de s’éroder et il a fini par tomber en disgrâce. D’une part, les biologistes ne s’entendent pas sur ce qu’est réellement un fossile vivant. Est-ce une même espèce qui n’a pas changé depuis sa première apparition dans les archives fossiles, ou est-ce le survivant d’une lignée autrefois florissante mais presque entièrement disparue depuis ? Un tel organisme doit-il être rare et peu répandu, ou peut-il être commun du moment que nous savons qu’il est issu d’une lignée ancienne ? Doit-il exister des vestiges fossiles ou suffit-il que son origine ancienne soit attestée par ses relations avec d’autres organismes ? Ou le fait de posséder des caractères primitifs (plésiomorphes) suffit-il à définir un fossile vivant ? Les séquences ADN présentes dans mes cellules, qui remontent au premier protiste unicellulaire respirant de l’oxygène, font-elles de moi un fossile vivant ?
Finalement, le concept s’est tellement dilué et a si souvent été appliqué de manière erronée qu’il a pratiquement perdu toute signification, tandis que les créationnistes, dans une manifestation typique de littéralisme infantile, se servent du terme comme preuve de l’absence d’évolution. La confusion est exacerbée par l’idée fausse que les organismes actuels ressemblant à des espèces éteintes sont les ancêtres encore vivants d’espèces modernes, ce qui n’est pas le cas. (Il existe bien des espèces résultant d’une hybridation et dont les deux espèces parentes vivent toujours mais c’est une autre histoire qui n’a rien à voir avec les fossiles vivants). On s’est également aperçu que certains fossiles vivants classiques (le tuatara de Nouvelle Zélande par exemple) évoluent au niveau moléculaire beaucoup plus vite que beaucoup d’organismes jeunes. Surtout, nous savons que tous les animaux et plantes considérées comme fossiles vivants sont des espèces modernes, récemment apparues en adaptation à l’environnement actuel, et non des survivants miraculeux génétiquement identiques aux vestiges pétrifiés, en dépit de ressemblances superficielles. Pour ces raisons, beaucoup de biologistes considèrent maintenant le terme de fossile vivant comme éteint.
Et pourtant, nous ne pouvons échapper au fait que certaines espèces actuelles ont une ascendance très ancienne, beaucoup plus vieille que les organismes modernes. Certaines sont les uniques survivants de lignées qui ont autrefois dominé les anciens écosystèmes, avant que ce fleuve d’espèces se réduise à un filet d’eau. Le cœlacanthe, représentant d’un groupe de poissons autrefois florissant, en est un. D’autres, comme les artémies, sont communs et appartiennent à des groupes encore riches d’espèces, – ici les crustacés – mais il en existe aussi des fossiles vieux d’un demi-milliard d’années. D’autres encore, comme les requins, au squelette cartilagineux, sont des organismes actuels présentant des caractères à la fois primitifs et modernes. Clairement, ces organismes doivent pouvoir nous apprendre des choses sur les mondes anciens.
Pour éviter ambiguïté du terme fossile vivant, les biologistes préfèrent appeler ces organismes reliques. Dès 1944, le paléontologue américain George G. Simpson a proposé un système de classification des espèces reliques reposant sur les circonstances ayant conduit à leur statut actuel. Certaines espèces sont devenues reliques à cause de la disparition de leur habitat, les confinant à de rares refuges écologiques, minuscules vestiges d’écosystèmes autrefois vastes (reliques écologiques). Mais la plupart des reliques sont issues de processus évolutifs, par exemple la concurrence d’une lignée apparentée plus efficace (reliques phylogénétiques), ou la fragmentation progressive et l’extinction subséquente à l’intérieur d’une grande partie de leur ancienne aire de distribution (reliques biogéographiques). Près de soixante-dix ans après son introduction, le système de Simpson n’a été que marginalement amélioré par les biologistes, preuve de sa validité.
Si les espèces reliques ne peuvent plus être considérées comme les répliques d’organismes vivant à des époques géologiques antérieures – ce sont en fait des organismes parfaitement adaptés aux conditions modernes – leurs morphologies et leurs comportements peuvent nous donner une idée de la vie au temps de leur gloire passée. Le comportement de ponte des limules en dit long sur les dangers de l’ère Mésozoïque qui les ont poussé à quitter l’eau pour la terre ferme, hostile, mais néanmoins préférable. La morphologie des ailes des Prophalangopsidés, parents de certains des premiers insectes chanteurs, nous donne un aperçu de l’origine de la production de sons chez ces animaux. La toxicité des cycas indique une pression adaptive forte de la part d’anciens herbivores, tandis que leur mode de pollinisation élucide les premiers chapitres de la longue relation entre les plantes et les insectes. L’aspect d’une touffe de prêles nous donne une idée des forêts du Crétacé, tandis que le dévouement maternel des blattes nous aide à comprendre l’évolution des sociétés complexes chez les insectes. Chacun de ces organismes reliques jette un modeste rayon de lumière sur un fragment de l’histoire de la vie et nous aide à mieux comprendre l’origine de la merveilleuse diversité des formes de vie, notre espèce comprise.
Les reliques peuvent aussi nous aider à comprendre pourquoi une espèce réussit mieux que les autres dans la quête sans fin pour transmette ses gênes. Il n’existe pas de caractère commun à tous les organismes reliques, mais des lignes directrices apparaissent. Certaines lignées ont survécu en s’adaptant aux conditions rudes de la haute montagne qui ont éliminé la plupart de leurs concurrents. D’autres doivent probablement leur longévité à des mécanismes de défense ingénieux et efficaces, tels que toxines puissantes ou substances chimiques indigestes. Certains groupes reliques ont su tirer parti d’autres organismes pour vivre. Parfois en les incorporant dans leurs propres tissus (les cycas et leurs symbiontes cyanobactériens en sont un bon exemple), parfois en profitant de nouvelles niches écologiques créées par des organismes plus récents et plus évolués comme les fougères qui ont occupé les nouveaux habitats créés par les plantes à fleur).Mais la stratégie la plus courante des reliques est simplement d’être des généralistes écologiques – le moyen le plus rapide de disparaître est d’être trop spécialisé. La constante des écosystèmes terrestres est d’évoluer en permanence ; si vous êtes trop dépendant d’un système précis de conditions, vous êtes en danger dès que l’inévitable changement survient.
[…] J’étais arrivé dans mon lieu de recueillement préféré, où s’ancre ma foi dans les forces physiques, indéniables et prouvées, qui ont donné naissance à notre univers. J’étais venu voir un spectacle, peut-être l’un des plus vieux de la planète, qui en est déjà à plus de cent millions de représentations.
Chaque année en mai et juin, pendant les quelques nuits qui coïncident avec la pleine et la nouvelle lune, les limules quittent les bancs de sable de l’océan Atlantique, et font irruption dans notre monde, sec et aussi étrange pour elles que leur domaine humide et sombre l’est pour nous. Luttant contre la traction des vagues refluantes, ces animaux extraordinaires marchaient lentement vers la plage. Les vagues s’écrasaient sur elles, les retournaient sur le dos et les ramenaient dans les eaux plus profondes, comme si l’océan ne voulait pas relâcher sa prise sur ses sujets. Et pourtant, elles persistaient. Lentement, mais sûrement, elles commençaient à émerger. Risquant leurs vies, elles abordaient un terrain étrange et inconnu où l’absence d’eau semblait démultiplier la force de la gravité. Dans l’eau, les limules sont étonnamment gracieuses, capables de courir sur le fond sableux et parfois même de nager maladroitement sur le dos. Mais ici, sur les plages de la baie du Delaware, elles se traînaient lentement. Les femelles étaient particulièrement désavantagées. Non seulement elles sont plus grandes et plus lourdes que les mâles, mais quand elles émergent sur le rivage, chacune a au moins un prétendant accroché à son dos. Parfois, elles traînent jusqu’à deux ou trois mâles soucieux de fertiliser les œufs qu’elles s’apprêtent à pondre.
Pendant que des centaines de mouches piqueuses faisaient tout leur possible pour nous vider de notre sang, mon ami et collègue Joe Warfel se tenait avec moi sur la plage en attendant que le spectacle commence. Le soleil s’obscurcissait et la marée haute d’approchait de son étale. Il y avait peu de gens sur la plage à notre arrivée, mais maintenant nous étions les seuls témoins de ce qui allait se passer.[…]
D’abord sont arrivées les grandes femelles. Presque toutes remorquaient des mâles. Dans la lumière déclinante, nous apercevions les queues épineuses de centaines de limules trébuchant dans les vagues pour gagner la terre ferme. Quand le soleil fut couché, la plage était recouverte de centaines d’énormes animaux luisants. Les femelles ont creusé le sable pour y déposer leurs œufs – près de 4 000 par nid -, tandis que les mâles se disputaient le privilège de les féconder. La fécondation étant externe, il arrive souvent que plusieurs mâles se partagent la paternité d’une même ponte. Dotés d’une paire de gros yeux composés (et huit autres plus petits) capables de voir la lumière ultraviolette, les mâles repèrent très bien les femelles dans le tourbillon des vagues de sable et de centaines d’autres mâles. Les scientifiques qui les étudient pensaient d’abord que les mâles étaient attirés par des phéromones femelles, mais il s’est avéré qu’ils se fient uniquement à leur excellente vue. Cela dit, ils font parfois des erreurs et il n’est pas rare de voir des chaînes de mâles qui se défont dès que les vraies femelles se montrent.
Assister au spectacle du frai massif des limules est pour moi ce qui se rapproche le plus d’une expérience religieuse. Mon cœur semble ralentir et un calme naturel m’aide momentanément à oublier les maux de ce monde. Aussi étranges et différentes de nous que les limules puissent paraître, ces créatures majestueuses me rappellent que nous partageons tous le même héritage évolutif. Bien que nos chemins aient divergé très tôt, les humains et les limules ont eu à un moment donné un ancêtre commun. C’était il y a très, très longtemps. Les limules sont sur Terre depuis plus longtemps que la plupart des lignées animales actuelles. Dans le dépôt fossile du Manitoba, au Canada, la découverte récente d’un petit animal nommé Lunataspis aurora prouve que des limules très semblables aux espèces modernes vivaient déjà à l’Ordovicien, il y a 445 millions d’années. Quand les dinosaures ont commencé à terroriser la terre au Trias, il y a 245 millions d’années, les limules étaient déjà des reliques d’ères géologiques passées. Et pourtant, elles ont survécu. Les dinosaures ont tiré leur révérence, la polarité et le climat de la Terre ont changé plusieurs fois, mais les limules ont poursuivi leur lent cheminement. Les espèces du Jurassique étaient si semblables aux actuelles que je doute que je remarquerais quoi que ce soit d’insolite si l’une d’elle rampait devant moi sur la plage du Delaware. Les limules paraissent avoir trouvé une morphologie et un mode de vie si réussis qu’elles ont survécu aux changements qui ont balayé des milliers de lignées en apparence plus imposantes (on pense aux dinosaures et aux trilobites). Contrairement à ce qu’affirment les créationnistes et autres lunatiques, elles ont continué d’évoluer. Les limules actuelles, qui se limitent à trois espèces en Asie du Sud-Est et une dans l’est de l’Amérique du Nord, diffèrent par bien des détails de leurs parents fossiles. Par exemple, nous savons que beaucoup et peut-être la plupart des limules fossiles vivaient en eau douce, souvent dans des marécages peu profonds surplombés par une végétation dense et certaines pourraient avoir été entièrement terrestres. Actuellement, seule la limule des mangroves (Carcinoscorpius rotundicauda) dans la péninsule malaise, fréquente régulièrement les rivières et pond en eau douce ou saumâtre.
Mais la morphologie des limules a changé très lentement. Une telle évolution morphologique très lente, appelée bradytélie, caractérise les organismes polyvalents. Être très spécialisé, par exemple se nourrir seulement d’herbes quand l’herbe est abondante, vous donne un grand avantage sur les organismes qui ne raffolent pas d’herbe. Mais que le climat change, remplaçant l’herbe par les cactus, et tous les herbivores exclusifs disparaissent alors que les généralistes s’en sortent. Les limules sont très tolérantes aux variations de l’environnement. L’espèce atlantique, Limulus Polyphemus, est présente dans les eaux glacées de la Nouvelle Ecosse, Canada, jusqu’aux littoraux tropicaux du Golfe du Mexique. Elle tolère de fortes variations de salinité, mange à peu près tout ce qui est organique (avec une préférence pour les moules et les coques)et peut survivre plusieurs jours hors de l’eau. C’est la signature d’un champion de la survie. La solide cuirasse des limules les met à l’abri de presque tout ennemi naturel (seuls les requins et les grandes tortues marines attaquent parfois les limules adultes). Mais la limule est une amoureuse, pas une guerrière. La longue pointe effrayante qui prolonge son corps n’est rien d’autre qu’un levier qui lui permet de se redresser lorsqu’une vague la retourne. Curieusement, cette queue appelée telson porte une rangée de cellules photosensibles, sortes d’yeux rudimentaires. Leur fonction probable est de les aider à s’orienter vers la plage au moment du frai.
Outre leur épais exosquelette, les limules asiatiques se défendent pendant la saison de reproduction à l’aide de toxines puissantes présentes dans les œufs et dans les parties molles du corps. La saxitoxine qu’elles contiennent est l’un des poisons organiques les plus puissants que l’on connaissance. Les premiers symptômes de défaillance musculaire apparaissent quelques minutes après l’injection. La mort par paralysie des muscles respiratoires intervient dans les 16 heures. Il n’existe pas d’antidote, seulement un traitement symptomatique qui peut laisser la victime avec des lésions cérébrales. Il est donc étrange de constater que ces limules figurent toujours au menu en Thaïlande et dans d’autres pays d’Asie. Il est connu qu’il ne faut pas manger ces animaux pendant la saison de reproduction, quand ils contiennent la toxine, mais une série de 280 empoisonnements par les limules dans un seul hôpital thaïlandais montre que le message n’est pas parvenu aux habitants de la ville de Chon Buri.
L’espèce américaine n’est pas toxique, mais les habitants doivent avoir un palais plus raffiné (ou moins ?) que les gourmets asiatiques, car manger les limules n’a jamais pris en Amérique du Nord. En 1588, un rapport de la première expédition en Virginie informait les Européens de l’existence de ces animaux et précisait que les indigènes en mangeaient, mais semble-t-il uniquement quand ils ne trouvaient rien de mieux. En Chine, la première mention écrite des limules date de 250 ap. J.C. Mais les auteurs ont dû seulement entendre parler du poisson hou et non le voir, car il était décrit comme une carpe à six pattes avec une grande nageoire dorsale. Il a fallu près de 1 400 ans pour avoir une description exacte. En 1666, l’auteur japonais Tekisai Nakamura a publié un dessin remarquablement précis de limule, le baptisant kabutogani (crabe cuirassé) – pas tout à fait juste, mais pas loin.
De son côté, le nom anglais de crabe fer-à-cheval jette la même confusion. Classés dans un groupe à part, les Xiphosures, les limules sont en effet plus proches des arachnides que des crabes et autres crustacés. Contrairement aux crabes, leur exosquelette ne contient pas beaucoup de calcium, et il est essentiellement fait de chitine, un polysaccharide. Solide, souple, biodégradable et non allergénique, la chitine de ces animaux est un matériau idéal pour les sutures chirurgicales, les implants et les lentilles de contact. Qui sait si vous ne regardez pas le monde à travers un morceau de limule ?
Le lendemain matin, Joe et moi avons trouvé la plage couverte d’œufs. Bien reposées et prêtes à commencer une nouvelle journée de festin, les mouches piqueuses nous ont attaqués avec un enthousiasme renouvelé. Tout en les écrasant par dizaines d’un coup, nous retournions les limules immobilisées sur le dos en recherchant des pontes particulièrement grosses. Bien que les femelles enfouissent leurs œufs dans le sable, la marée en découvre beaucoup. Les œufs tout juste pondus ne sont pas plus gros qu’un grain de riz. Étonnamment, ils sont deux fois plus gros à la fin de leur développement. Ce qui est bien sûr impossible. Leur croissance est une illusion qui résulte de la production d’une mince membrane externe par l’embryon. Un œuf pleinement développé, après deux semaines dans le sable, ressemble à un petit bocal renfermant une petite limule, impatiente de briser les murs de sa prison. Une fois libres, les larves (du moins les chanceuses) profitent d’une vague pour retourner à l’océan, où elles flotteront librement pendant une semaine avant de rejoindre le fond dans les eaux littorales peu profondes, pour commencer une vie identique à celle de leurs parents.
Mais une question demeure : pourquoi des animaux aquatiques quittent-ils leur milieu naturel pour exposer leurs petits aux dangers de la terre ferme, quand leur développement nécessite de l’eau ? Il serait tentant de comparer ce comportement à celui des tortues marines, qui passent aussi leur vie dans l’océan et ne le quittent que pour pondre dans le sable. Mais les raisons de ces comportements similaires ne pourraient pas être plus différentes. Descendant de reptiles terrestres, les tortues marines sont des nouvelles venues dans l’océan et doivent respirer à la surface. Elles ne peuvent pas rester sous l’eau plus de deux heures (au repos, moins si elles sont actives), et leurs œufs doivent être exposés à l’oxygène atmosphérique pour éviter l’asphyxie des embryons. Inversement, les limules respirent sous l’eau et leurs œufs doivent être submergés ou au moins fréquemment recouverts par les vagues pour se développer. La force qui les a poussées à une mesure aussi désespérée, ce sont les millions d’estomacs affamés qui attendaient dans l’océan la provende des œufs de limules. Lorsque les limules sont apparues sous leur forme actuelle, il y a 445 millions d’années, presque toute la vie animale – et quasiment tous les prédateurs – vivaient dans les mers et les lacs du globe. En osant sortir de leur domaine aquatique, les limules ont donné à leur progéniture un avantage considérable sur les autres organismes marins. Elles ont été parmi les premiers animaux, sinon les tout premiers, à adopter une stratégie d’exportation pour leur reproduction. Aucun animal terrestre actuel ne descend des limules, mais il est vraisemblable que certains des premiers invertébrés terrestres aient suivi une voie similaire, poussés par la pression sélective des prédateurs d’œufs.
Mais il y a une chose que les limules ne pouvaient pas prévoir. Les dinosaures, ces tétrapodes gigantesques qui ont foulé la terre pendant presque 170 millions d’années avant de sembler s’éteindre à la fin du Crétacé, n’ont pas complètement disparu. Certains ont évolué pour donner une des lignées les plus prospères du règne animal – les oiseaux -. Les oiseaux sont malins et ont une excellente vision. Ils ont aussi un métabolisme intense qui leur permet de maintenir une température corporelle constante, mais qui nécessite un apport continu d’aliments énergétiques. Les œufs de limule sont exactement ce qu’il leur faut.
Bourrés de lipides et de protéines, les œufs de limules atlantiques sont un carburant idéal pour de nombreux oiseaux du littoral. Avec la régularité d’un coucou suisse, les limules répandent un déjeuner d’œufs frais sur les rivages de la baie du Delaware aux pleines et nouvelles lunes de la fin du printemps. Un oiseau, le terne bécasseau maubèche (Calidris canutus rufa) doit même sa survie aux limules. Sa migration inclut un arrêt dans la baie du Delaware répit indispensable après un voyage non-stop depuis l’Amérique du Sud. Les nuages de bécasseau s’abattant sur les plages de la baie au printemps ont toujours stupéfié les hommes. Mais, il y a quelques années, quand la population de limules a chuté à une fraction de sa splendeur passée, le bécasseau s’est mis à disparaître à son tour.
Les ornithologues américains se sont émus. Des pétitions ont circulé, des études ont été réalisées et des lois protégeant ces oiseaux ont été votées. Quelqu’un a fini par comprendre que si les limules disparaissaient, le précieux oiseau suivrait ; il faudrait peut-être aussi sauver ce méprisable invertébré. Au New Jersey, un an avant d’écrire ces lignes, on pouvait sans problème remplir son pick-up de centaines de limules pour servir d’appât ou autre. Maintenant, il est illégal d’en ramasser une seule. J’ai été menacé d’une amende de 10 000 $ pour avoir ramassé une limule (dans l’intention de la relâcher aussitôt après avoir pris quelques photos) par un garde particulièrement zélé. Il m’a donné un avertissement sévère mais il m’a fait grâce. En repartant, il a marché sur une des dizaines de limules qui gisaient sur le dos. Il ne s’est pas arrêté.
Il y a un siècle environ, nous n’aurions probablement pas pu arpenter la plage sans marcher sur les limules. Elles étaient si nombreuses que les hommes ont cherché des moyens d’utiliser cette abondante ressource. L’idée a été vite trouvée. De 1880 à 1920, on a ramassé plus d’un million de limules par an pour les tuer et les broyer afin d’en faire de l’engrais ou de l’aliment pour cochons. Cette pratique a continué jusque dans les années 1970, lorsque la dernière usine a fermé, en grande partie à cause des plaintes motivées par l’odeur, mais aussi parce que la récolte était tombée à 100 000 par an. Mais une autre industrie l’a remplacée, tuant les limules pour appâter les anguilles et les bulots. Tout ceci a entraîné un déclin de la population de limules dont elle pourrait ne jamais se remettre complètement. Et bien sûr, il y a la question du sang de limule.
Au début des années 1950, le chercheur Frederik Bang a découvert que le sang de ces animaux était extrêmement sensible aux endotoxines produites par les bactéries Gramnégatives. Toute exposition du système immunitaire de la limule à ces bactéries provoque une coagulation généralisée et immédiate, isolant efficacement les microbes et les empêchant de nuire. Bang et son collaborateur Jack Levin ont rapidement réalisé les possibilités offertes par cette découverte. Après avoir isolé les éléments actifs du sang de limule, les amébocytes, ils ont mis au point un extrait appelé lysat d’amébocyte de limule (LAL). Celui-ci facilite la détection des endotoxines bactériennes dans l’urine, le liquide céphalo-rachidien et autres fluides corporels. Mais sa principale application est la détection des contaminations dans les médicaments intraveineux et sur les instruments médicaux. Si on vous a fait une injection intraveineuse ou si on vous a remplacé une valve cardiaque, leur stérilité a très probablement été testée avec du LAL. La récolte des limules pour produire du LAL est maintenant une industrie importante qui génère des centaines de millions de $ par an. Chaque année, on capture ces centaines de milliers de limules pour prélever 20 % de leur sang bleu (il contient de l’hémocyanine à base de cuivre au lieu d’hémoglobine à base de fer). Les animaux sont ensuite relâchés en mer. Bien que la mortalité des limules relâchées soit faible (plus de 85 % des animaux survivent à ce prélèvement de sang), l’industrie perturbe la structure de leurs populations en déplaçant ces animaux à des centaines de kilomètres des lieux de capture.
Ce qui me choque le plus, c’est qu’il a fallu un oiseau pour que l’on commence à s’inquiéter du déclin des limules. Celles-ci ont été systématiquement exterminées pendant plus d’un siècle dans ce que je ne peux qu’appeler un phylogénocide. Cela peut sembler sévère mais réfléchissez-y : la perte d’un bécasseau maubèche, aussi impardonnable qu’elle soit, ne signifie que la perte d’un dix millième du patrimoine génétique des oiseaux. La perte d’une espèce de limule signifie la perte d’un quart de l’héritage génétique des Xiphosures, l’une des lignées les plus anciennes de la planète. Et pourtant, nous nous préoccupons plus d’une adorable petite boule de plumes, dont notre bien-être dépend peu, que d’une bestiole répugnante qui a déjà sauvé des millions de vies humaines. Comme nous sommes superficiels…
L’extinction des limules est presque complète au Japon, où l’espèce locale (Tachypleus tridentatus) était aussi abondante que sa cousine atlantique. J’y suis allé à l’été 2008 pour l’observer sur la dernière plage japonaise où elles se montrent encore en grand nombre. Avec mon ami Kenji Nishida, entomologiste et grand photographe, je me suis rendu sur la plage d’Imari, sur l’île de Kyushu. C’était un jour avant le festival Kabotugani, une célébration annuelle des limules. On nous a raconté que c’était une bonne année : quatre couples de limules avaient été aperçus près de la plage ! Quatre couples. Huit individus. C’était tout. Dans les années 1980, il n’était pas rare de voir cinq cents limules au même endroit. Au Japon, où les limules sont presque révérées, l’Association Nippone pour la Protection des Limules se bat pour la survie de l’espèce. Pourtant, leur nombre continue à décroître. Il y a là une leçon à apprendre. Dans le destin d’une espèce, il existe un point de non-retour. Espérons que nous pourrons empêcher la limule atlantique de l’atteindre.
Chaque fois que je rentre de la baie du Delaware, je me demande ce que je trouverai l’année suivante. Est-ce mon imagination, ou y-a-t-il de moins en moins de limules chaque année ? Par chance, cela ne semble pas être le cas. Les efforts de protection ont des effets positifs qui, espérons-le, arrêteront le déclin de ces animaux magnifiques. Dans la baie, on a instauré une zone d’interdiction de ramassage, la réserve Carl N. Schuster, d’après l’un des plus éminents chercheurs et avocats des limules. La récolte des limules pour appâts est limitée et les gens s’intéressent de plus en plus au sort des limules. Les volontaires sont chaque année plus nombreux pour dénombrer les limules au moment du frai. Récemment, j’ai vu dans un magasin de jouets, à côté d’un panda et d’une orque, un limule en peluche. Cela signifie-t-il que maintenant les enfants trouvent les limules adorables ? Peut-être y-a-t-il de l’espoir finalement.
Piotr Naskrecki. Reliques. Ulmer 2012
ère géologique : Silurien : 443.4 à 419 millions d’années.
440 m.a.
Des explosions astrophysiques, appelées sursauts gamma, sous-produit soit de supernovae, soit de collisions entre étoiles ultradense nommées neutroniques, pourraient avoir provoqué des extinctions d’espèces et un refroidissement du climat : destruction d’une bonne partie de la couche d’ozone, conversion de l’azote et de l’oxygène de l’air en dioxyde d’azote : il n’en faut pas plus pour perturber gravement la vie.
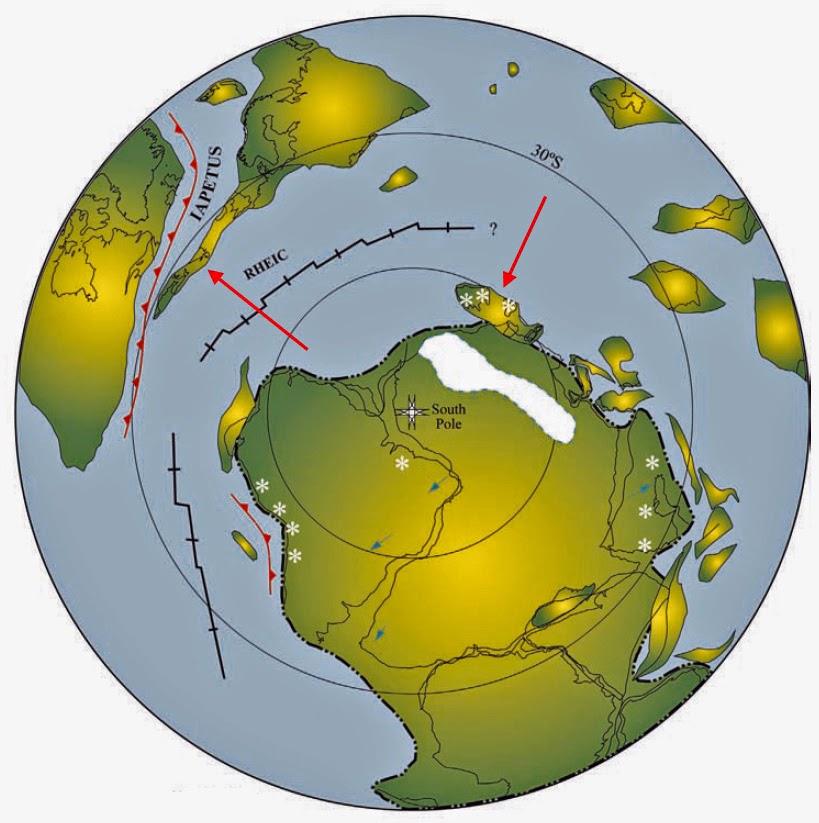
Dernière reconstruction de l’hémisphère sud à l’ordovicien. Dans le dernier silurien ordovicien-ancien (440 Ma), Avalonia (ouest et est situés à la flèche de gauche) s’était précédemment écartée de la marge nord du Gondwana, à la dérive de la fermeture d’Iapetus Ocean et se préparait bientôt à entrer en collision avec Baltica et Laurentia au cours de l’orogenèse Acadian-Caledonide. Le corps principal du Gondwana polaire sud est en train de traverser un océan Rheic en déclin. Les terranes de type Cadomian (flèche vers la droite) sont toujours amarrés en marge de Gondwana.
de 445.2 m.a. à 443.8 m.a.
Un astéroïde aurait percuté l’Australie, alors partie du Gondwana, créant un cratère de 520 km Ø : Deniliquin, le nom de la ville la plus proche, au sud-est de l’Australie, en Nouvelles Galles du Sud, au nord de Melbourne et à l’est de Victor Harbor, près d’Adélaïde. Ce sont des chercheurs de l’Université de Nouvelle-Galles du Sud, à Sydney, qui l’ont découvert en 2023… alors qu’ils cherchaient de l’or !
Les astéroïdes ont joué un rôle crucial dans la formation et l’évolution de la Terre. Durant des milliards d’années, leurs impacts ont façonné la géologie, influencé le climat et même provoqué des extinctions massives. Ces événements cataclysmiques ont laissé des empreintes indélébiles, mais souvent cachées sous des couches de roches ou des océans. De plus, ces dernières peuvent s’éroder, rendant difficile leur identification.
Au cœur de l’Australie du Sud-Est, une gigantesque structure enfouie vient d’être dévoilée. Son étude, menée par des experts de l’UNSW (Université de Nouvelle-Galles du Sud, à Sydney), met en lumière ce qui pourrait être la plus grande structure d’impact d’astéroïde jamais découverte. Au-delà de sa taille impressionnante, elle offre une perspective inédite sur les conséquences géologiques des impacts et leur rôle dans l’évolution de notre planète. L’étude est publiée dans la revue Tectonophysics.
En complément de ces motifs magnétiques, le cratère présente des motifs gravitaires circulaires de Bouguer. Ces motifs mettent en évidence des variations de la gravité à la surface de la Terre, souvent utilisées pour identifier des structures souterraines ou des variations de densité dans la croûte terrestre.
Les mesures magnétiques montrent également des preuves de failles radiales : des fractures qui rayonnent à partir du centre d’une grande structure d’impact. Cela s’accompagne en outre de petites anomalies magnétiques qui peuvent représenter des digues ignées, qui sont des feuilles de magma éjectées dans des fractures au sein d’un corps rocheux préexistant.

Les auteurs soulignent également dans un article qu’ils ont publié dans The Conversation que la taille de cette structure est tout simplement stupéfiante. Avec un rayon d’environ 260 km, elle s’étend sur une vaste zone, témoignant de l’ampleur de l’événement qui a pu la créer. Cette dimension la place parmi les plus grandes structures géophysiques de ce type découvertes à ce jour.
La caractéristique de Deniliquin, malgré sa position profondément enfouie sous la surface, révèle des indices intrigants d’un événement passé. Lorsque les chercheurs ont effectué des forages dans cette région, ils ont découvert des lamelles de Boehm. Ces formations, qui sont des minces couches ou inclusions dans des minéraux, peuvent être indicatives de processus géologiques spécifiques.

Néanmoins, d’autres éléments pointent vers un impact d’astéroïde. La structure à anneaux multiples est atypique et ne correspond pas aux formations géologiques régionales environnantes. De plus, la présence d’une zone magnétique centrale silencieuse est un autre indice fort. Ces caractéristiques, combinées, suggèrent que nous sommes en présence de la zone racinaire, c’est-à-dire la partie la plus profonde, d’une vaste structure d’impact. Cette zone racinaire serait le vestige direct de l’endroit où l’astéroïde a frappé, ayant depuis été recouverte par des couches de roche et de sédiments au fil du temps.
De plus, cette découverte pourrait servir de point de départ pour de nouvelles recherches. Les scientifiques pourraient être en mesure d’étudier en détail les conséquences d’un tel impact sur l’environnement local et global. Cela inclut les changements climatiques potentiels, les perturbations de l’écosystème, et même les extinctions qui pourraient avoir été déclenchées par l’impact.
Selon les auteurs, celui-ci aurait pu se produire lors de l’événement d’extinction massive de l’Ordovicien supérieur. Ils précisent : Plus précisément, je pense que cela a peut-être déclenché ce qu’on appelle le stade de glaciation hirnantien, qui a duré entre 445,2 et 443,8 millions d’années, et est également défini comme l’événement d’extinction ordovicien-silurien. 85% des espèces de la planète ont disparu à cette époque, soit le double de l’impact de Chixculub, qui a tué les dinosaures.
Trust my science
430 m.a.
Jusqu’alors, l’alimentation en eau des plantes se faisaient par capillarité, chaque paroi cellulaire se gorgeant d’eau, mais le système n’est efficace que jusqu’à une hauteur de 2 cm, ce qui limite radicalement la croissance verticale ; vont apparaître des cellules lignifiées dont la mort programmée permet la transformation en canal de transport de l’eau : ces larges cellules mortes et vides étaient un million de fois plus efficaces que l’ancien mécanisme intercellulaire, permettant le transport sur de plus longues distances et un taux de diffusion du CO2 plus élevé. Cette tige unique est porteuse du sporange, l’organe reproducteur de spores : spores et oosphères n’ont dès lors plus besoin d’eau pour se disperser : le vent y pourvoie. En se verticalisant, la plante développe un système de vaisseaux pour diffuser, avec la sève, sucre et matière organique dans tout leur organisme.
Les plantes ayant acquis ce système de transport étaient devenues capables d’extraire de l’eau de leur environnement par des organes semblables à des racines, et cette dépendance moindre aux aléas climatiques leur permettait d’atteindre des tailles bien plus importantes. Mais, conséquence de cette relative indépendance vis-à-vis de l’environnement, elles perdirent leur capacité à survivre à la dessiccation, un trait trop coûteux à conserver.
ère géologique : Dévonien : 419 à 358.9 millions d’années
416 m.a.
La concentration atmosphérique en O2 dépasse 13 % : dès lors les incendies deviennent possibles : on en a les premières traces fossiles, enchâssés dans du charbon.
__________________________________________________________________________________________________
[1] On peut en trouver dans des environnements tout à fait imprévus… jusqu’à 2000 mètres sous terre…
[2] Être vivant en symbiose étroite avec un autre être
[3] Corpuscules des cellules végétales porteuses de chlorophylle et sièges de la photosynthèse
[4] Organites composant les cellules et productrices d’énergie
[5] Et que dire encore de ces poteaux d’enceinte de cases, voire de ces poteaux télégraphiques, qui repartent en végétation en Afrique tropicale et équatoriale, quelques semaines après avoir été posés … et des ces bambous coupés, posés sur leur longueur sur du sable où l’eau n’est pas loin et qui repartent eux aussi en végétation par des racines développées au niveau des nœuds.